| . |
|
||||||
|
|
| . |
|
||||||
|
|
Les
membranes
plasmiques (comme les autres membranes biologiques),
sont des barri├©res s├®lectives qui r├®gulent le passage des substances
├Ā l'int├®rieur et ├Ā l'ext├®rieur des cellules. Le
transport membranaire
est le mouvement de mol├®cules ├Ā travers
ces membranes cellulaires. Ce processus intervient dans l'absorption des
nutriments par les cellules, l'├®limination de
leurs d├®chets, la communication cellulaire et le maintien de l'hom├®ostasie
cellulaire. Il existe deux principaux types de transport membranaire :
le transport passif, qui ne consomme pas d'├®nergie produite par la cellule,
et le transport actif qui en utilise, le plus souvent en faisant intervenir
l'hydrolyse de l'ATP (ad├®nosine
triphosphate).
Transport passifLe moyen le plus simple pour une cellule d'├®changer des substances avec son environnement ├Ā travers la membrane plasmique est celui qui ne requiert aucune consommation d'├®nergie par la cellule. L'├®nergie en jeu est l'├®nergie cin├®tique (dont l'expression macroscopique est la temp├®rature) des atomes, ions et mol├®cules. On parle alors de transport passif.Dans le transport passif, les substances passent d'une zone de concentration plus ├®lev├®e ├Ā une zone de concentration plus faible au cours d'un processus appel├® diffusion. On parle de gradient de concentration pour d├®crire un espace physique dans lequel il existe entre un point et un autre une variation de concentration d'une seule substance. Lorsque la travers├®e de la membrane plasmique est possible (principe de la perm├®abilit├® s├®lective), trois m├®canismes de diffusion s'observent dans le transport passif : la diffusion simple, la diffusion facilit├®e et l'osmose. Perm├®abilit├®
s├®lective.
Les membranes plasmiques ont des r├®gions hydrophiles et des r├®gions hydrophobes. Cette caract├®ristique facilite le mouvement de certains mat├®riaux ├Ā travers la membrane et entrave le mouvement d'autres. Un mat├®riau liposoluble peut facilement glisser ├Ā travers le noyau lipidique hydrophobe de la membrane. Des substances telles que les vitamines liposolubles A, D, E et K passent ainsi ais├®ment ├Ā travers les membranes plasmiques du tube digestif et d'autres tissus. Les m├®dicaments liposolubles p├®n├©trent ├®galement facilement dans les cellules et sont facilement transport├®s dans les tissus et les organes du corps. Les mol├®cules d'oxyg├©ne et de dioxyde de carbone n'ont pas de charge et passent eux aussi par simple diffusion. A l'exception de l'eau, les substances polaires, rencontrent des difficult├®s pour traverser la membrane plasmique. Bien que certaines mol├®cules polaires se connectent facilement ├Ā l'ext├®rieur d'une cellule, elles ne peuvent pas passer facilement ├Ā travers le noyau lipidique de la membrane. De plus, alors que les petits ions pourraient facilement glisser ├Ā travers les espaces de la mosa├»que de la membrane, leur charge les en emp├¬che. Les ions tels que le sodium, le potassium, le calcium et le chlorure doivent avoir un moyen sp├®cial de passer au travers des membranes plasmiques. Les sucres simples et les acides amin├®s, parce que ce sont des substances insolubles dans les lipides ou parce que leurs mol├®cules sont de trop grandes dimensions, ont ├®galement besoin d'aide pour traverser ces membranes. La diffusion simple.
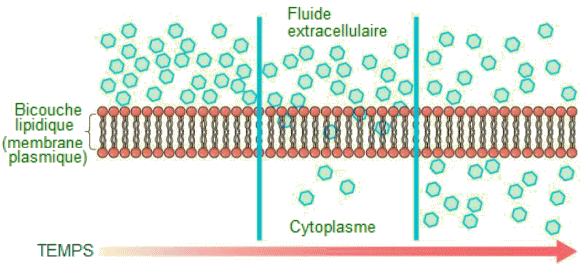
Dans un milieu donn├®, chaque substance, par exemple le fluide extracellulaire, a son propre gradient de concentration, ind├®pendant des gradients de concentration d'autres mat├®riaux. Chaque substance diffuse en fonction de ce gradient. Plusieurs facteurs affectent le taux de diffusion. ŌĆó L'├®tendue du gradient de concentration : plus la diff├®rence de concentration est grande, plus la diffusion est rapide. Plus la distribution du mat├®riau se rapproche de l'├®quilibre, plus le taux de diffusion est lent.Transport facilit├®. Dans le transport facilit├®, ├®galement appel├® diffusion facilit├®e, le mat├®riau se d├®place ├Ā travers la membrane plasmique gr├óce au concours de prot├®ines transmembranaires vers le bas d'un gradient de concentration (des points de haute concentration aux points de basse concentration). Ici encore sans d├®pense d'├®nergie cellulaire. Les substances qui b├®n├®ficient d'un transport facilit├® ne se diffuseraient pas facilement ou rapidement ├Ā travers la membrane plasmique. La solution pour d├®placer les substances polaires et d'autres substances ├Ā travers la membrane plasmique r├®side dans l'action des prot├®ines qui en couvrent la surface. 1) Le mat├®riau transport├® est d'abord attach├® aux r├®cepteurs de prot├®ines ou de glycoprot├®ines sur la surface ext├®rieure de la membrane plasmique. Cela permet au mat├®riau dont la cellule a besoin d'├¬tre retir├® du fluide extracellulaire.L'union entre les prot├®ines de transport et les mol├®cules transport├®es est transitoire.. Apr├©s le passage d'une mol├®cule transport├®e de l'autre c├┤t├® de la membrane, elle se d├®tache et la prot├®ine de transport ainsi lib├®r├®e peut entamer un nouveau cycle et permettre ├Ā de nouvelles mol├®cules de faire la travers├®e. Osmose.
Les cellules n'ont
aucun contr├┤le sur l'osmose. La direction dans laquelle circule l'eau
est seulement fonction de la concentration en mol├®cules d'eau ├Ā l'ext├®rieur
et ├Ā l'int├®rieur de la cellule. La plupart des membranes plasmiques sont
tr├©s perm├®ables ├Ā l'eau.
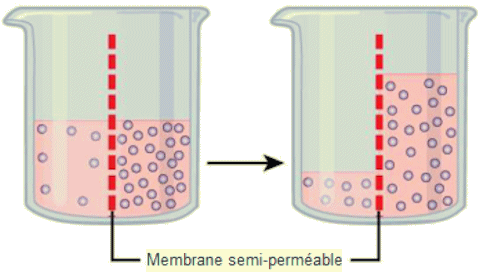
Le principe de la diffusion, on l'a dit, est que les mol├®cules se d├®placent et se r├®partissent uniform├®ment dans le milieu si elles le peuvent. Cependant, seul le mat├®riau capable de traverser la membrane diffusera ├Ā travers elle. Dans le cas de l'osmose, le solut├® ne peut pas diffuser ├Ā travers la membrane, mais l'eau le peut. Cette diffusion de l'eau ├Ā travers la membrane, l'osmose, se poursuivra jusqu'├Ā ce que le gradient de concentration de l'eau atteigne z├®ro. L'osmose se produit constamment dans les syst├©mes vivants. La tonicit├® d├®crit la quantit├® de solut├® dans une solution. La mesure de la tonicit├® d'une solution, ou la quantit├® totale de solut├®s dissous dans une quantit├® sp├®cifique de solution est appel├®e son osmolarit├® ou concentration osmotique. Trois termes: hypotonique, isotonique et hypertonique sont utilis├®s pour relier l'osmolarit├® d'une cellule ├Ā l'osmolarit├® du liquide extracellulaire qui contient les cellules. ŌĆó Solutions hypotoniques. - Dans une situation hypotonique, le liquide extracellulaire (par exemple l'eau du robinet) est moins osmolaire que le liquide ├Ā l'int├®rieur de la cellule et l'eau p├®n├©tre dans la cellule. Dans les syst├©mes vivants, le point de r├®f├®rence est toujours le cytoplasme, donc le pr├®fixe hypo- signifie que le liquide extracellulaire pr├®sente une concentration plus faible de solut├®s, ou une osmolarit├® inf├®rieure, que le cytoplasme cellulaire. Cela signifie ├®galement que le liquide extracellulaire a une concentration d'eau plus ├®lev├®e dans la solution que la cellule. Dans cette situation, l'eau suivra son gradient de concentration et entrera donc dans la cellule. 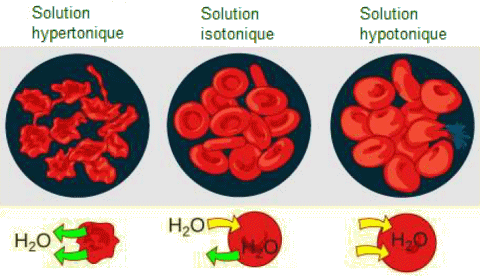
Transport actifLe transport actif par transporteur est un processus au cours duquel des substances sp├®cifiques sont transport├®es ├Ā travers la membrane cellulaire contre leur gradient de concentration ou un gradient ├®lectrochimique en utilisant des prot├®ines de transport sp├®cialis├®es. Ce mode de transport n├®cessite de l'├®nergie, g├®n├®ralement fournie par l'hydrolyse de l'ATP ou par l'exploitation d'un gradient ├®lectrochimique.Les m├®canismes de transport actif, contrairement ├Ā ceux impliqu├®s dans le transport passif, n├®cessitent l'utilisation de l'├®nergie de la cellule, g├®n├®ralement fournie par la mol├®cule d'ad├®nosine triphosphate (ATP). Ils concernent le passage de substances ├Ā travers la membrane plasmique dans le sens oppos├® au gradient de concentration, c'est-├Ā-dire d'une r├®gion ├Ā basse concentration vers une r├®gion plus concentr├®e. Certains m├®canismes de transport actif d├®placent ├Ā travers la membrane des mat├®riaux de faible poids mol├®culaire, tels que des ions. Les cellules doivent ├®galement pouvoir ├®liminer et absorber les mol├®cules et particules plus grosses. Certaines cellules sont m├¬me capables d'engloutir des micro-organismes unicellulaires entiers. Comme d'autres processus de transport, le transport actif par transporteur peut atteindre une saturation lorsque toutes les prot├®ines de transport sont occup├®es par des substrats. De plus, les transporteurs peuvent ├¬tre sp├®cifiques ├Ā certains substrats, ce qui signifie qu'ils ne transportent que des mol├®cules particuli├©res. Le transport actif par transporteur peut ├¬tre r├®gul├® par la modulation de l'activit├® des prot├®ines de transport ou par la r├®gulation de leur expression g├®nique, selon les besoins cellulaires. Exemple de transporteur : le transporteur de glucose, qui est responsable de l'absorption du glucose dans de nombreuses cellules. On peut donner comme exemple de transport actif le passage de l'iode du sang dans la glande thyro├»de. Le glucose, les acides amin├®s et certains ions passent aussi dans les cellules par ce type de transport. Gradient ├®lectrochimique.
L'int├®rieur des cellules vivantes est ├®lectriquement n├®gatif par rapport au fluide extracellulaire dans lequel elles baignent; en m├¬me temps, les cellules ont des concentrations de potassium (K+) plus ├®lev├®es et des concentrations de sodium (Na+) plus faibles que le liquide extracellulaire. Ainsi, dans une cellule vivante, le gradient de concentration de Na+ a tendance ├Ā le conduire dans la cellule, et le gradient ├®lectrique de Na+ (un ion positif) a ├®galement tendance ├Ā le conduire vers l'int├®rieur charg├® n├®gativement. La situation est cependant plus complexe pour d'autres ├®l├®ments comme le potassium. Le gradient ├®lectrique de K+, un ion positif, a ├®galement tendance ├Ā le conduire dans la cellule, mais le gradient de concentration de K+ a tendance ├Ā chasser K+ hors de la cellule. Le gradient combin├® qui affecte un ion
est appel├® son gradient ├®lectrochimique; il est particuli├©rement
important pour les cellules musculaires et nerveuses.
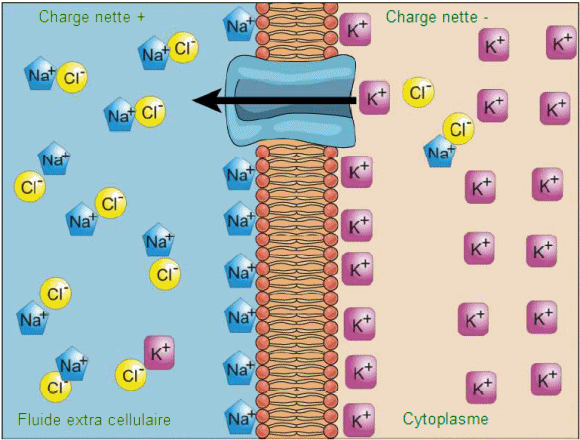
Transport ├Ā contre-gradient.
Les m├®canismes de transport actifs, aussibie que les prot├®ines qui sont ├Ā l'oeuvre, sont appel├®s collectivement pompes, pompes ioniques ou prot├®ines porteuses. Chaque pompe a ses fonctions diff├®rentes et ses substrats, mais elles partagent plusieurs caract├®ristiques et points communs dans leur fonctionnement et leur structure La plupart des pompes membranaires fonctionnent en utilisant de l'├®nergie d├®riv├®e de l'hydrolyse de l'ad├®nosine triphosphate (ATP), qui une source d'├®nergie cellulaire. L'├®nergie lib├®r├®e lors de cette r├®action est utilis├®e pour alimenter le transport actif des ions ou des mol├®cules ├Ā travers la membrane. Les pompes ioniques effectuent g├®n├®ralement un transport actif, ce qui signifie qu'elles d├®placent les ions ou les mol├®cules ├Ā travers la membrane contre leur gradient de concentration ou ├®lectrochimique. Cela n├®cessite l'utilisation d'├®nergie pour vaincre cette opposition et maintenir des gradients de concentration sp├®cifiques. Le fonctionnement des pompes implique souvent des changements conformationnels dans leur structure prot├®ique. Ces changements sont g├®n├®ralement d├®clench├®s par la liaison de substrats ou de co-facteurs. Les pompes fonctionnent g├®n├®ralement de mani├©re cyclique, alternant entre diff├®rentes ├®tapes telles que la liaison du substrat, la phosphorylation, le transport du substrat ├Ā travers la membrane, et la d├®phosphorylation pour permettre la r├®cup├®ration de la conformation initiale de la pompe. Les pompes membranaires sont souvent r├®gul├®es par divers m├®canismes (modifications post-traductionnelles, interactions avec d'autres prot├®ines ou des ligands, changements dans l'activit├® de la pompe en r├®ponse ├Ā des signaux cellulaires ou environnementaux). Enfin, les diff├®rentes pompes membranaires sont localis├®es dans des compartiments cellulaires sp├®cifiques ou dans des r├®gions sp├®cifiques de la membrane cellulaire, en fonction de leurs fonctions biologiques et de leurs besoins en termes de r├®gulation du transport ionique ou mol├®culaire. Pompe
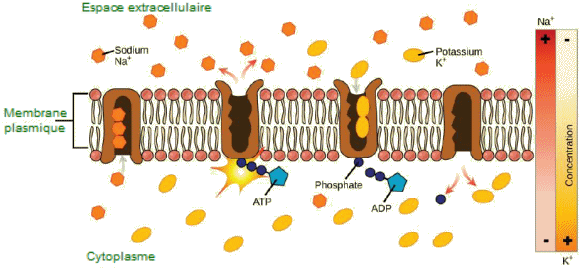
sodium-potassium.
La pompe est constitu├®e
de deux parties principales : une partie extracellulaire, o├╣ les ions
sodium (Na+) se lient, et une partie intracellulaire,
o├╣ les ions potassium (K+) se lient. Au
repos, la pompe a trois sites de liaison pour les ions sodium ├Ā l'ext├®rieur
de la cellule. Trois ions sodium se lient ├Ā ces sites ce qui d├®clenche
une modification de la forme de la pompe, entraînant ainsi la phosphorylation
d'une partie de la pompe par l'ATP (ad├®nosine triphosphate), transf├®rant
alors un groupement phosphate sur la pompe elle-même. Ce changement de
conformation favorise la lib├®ration des ions sodium vers l'ext├®rieur
de la cellule. En m├¬me temps, la pompe a une affinit├® accrue pour
les ions potassium ├Ā l'int├®rieur de la cellule. Deux ions potassium se
lient aux sites de liaison intracellulaires. La liaison des ions potassium
stimule la d├®phosphorylation de la pompe, ce qui la ram├©ne ├Ā sa conformation
initiale. Ce retour ├Ā la conformation initiale favorise la lib├®ration
des ions potassium ├Ā l'int├®rieur de la cellule. Au final, le processus
permet le maintien du gradient ├®lectrochimique, avec une concentration
plus ├®lev├®e de potassium ├Ā l'int├®rieur de la cellule et une concentration
plus ├®lev├®e de sodium ├Ā l'ext├®rieur.
Pompe
├Ā calcium.
Lorsqu'il y a une augmentation de la concentration de calcium ├Ā l'int├®rieur de la cellule, le calcium se lie ├Ā des sites sp├®cifiques sur la pompe calcium ATPase situ├®s ├Ā la face cytoplasmique (intracellulaire) de la membrane cellulaire. La liaison du calcium induit un changement conformationnel dans la pompe, ce qui permet ├Ā une partie de la prot├®ine d'effectuer une phosphorylation. Cela se produit gr├óce ├Ā l'utilisation de l'ATP, o├╣ un groupement phosphate est transf├®r├® ├Ā la pompe par une enzyme kinase associ├®e. Le processus de phosphorylation favorise le rel├óchement du calcium vers l'ext├®rieur de la cellule ou vers les organites intracellulaires tels que le r├®ticulum endoplasmique ou les mitochondries, selon le type de pompe calcium ATPase sp├®cifique. Ce mouvement de calcium est contre son gradient de concentration, n├®cessitant donc de l'├®nergie fournie par l'hydrolyse de l'ATP. Apr├©s le transport du calcium, la pompe reprend sa conformation initiale gr├óce ├Ā la d├®phosphorylation, qui est catalys├®e par une enzyme phosphatase associ├®e. Ce processus permet de pr├®parer la pompe pour de nouveaux cycles de liaison de calcium et de transport. sation cellulaire, la contraction musculaire et la s├®cr├®tion de neurotransmetteurs. Pompe
├Ā protons.
Au repos, la pompe ├Ā protons est li├®e ├Ā une mol├®cule d'ATP. Les protons se lient ├Ā des sites sp├®cifiques sur la prot├®ine, g├®n├®ralement situ├®s du c├┤t├® cytoplasmique (intracellulaire) de la membrane. La liaison des protons induit un changement conformationnel dans la pompe, ce qui permet ├Ā une partie de la prot├®ine d'effectuer une phosphorylation. Le processus de phosphorylation favorise le transport des protons ├Ā travers la membrane, contre leur gradient ├®lectrochimique. La pompe travaille ainsi activement pour accumuler les protons de part et d'autre de la membrane, cr├®ant ainsi un gradient de concentration et un potentiel ├®lectrique. Apr├©s le transport des protons, la pompe reprend sa conformation initiale gr├óce ├Ā la d├®phosphorylation, qui est catalys├®e par une enzyme phosphatase associ├®e. Ce processus permet de pr├®parer la pompe pour de nouveaux cycles de liaison de protons et de transport. Pompe
├Ā chlorure.
Au repos, la pompe ├Ā chlorure est li├®e ├Ā une mol├®cule d'ad├®nosine triphosphate (ATP). Les ions chlorure se lient ├Ā des sites sp├®cifiques sur la prot├®ine, g├®n├®ralement situ├®s du c├┤t├® cytoplasmique (intracellulaire) de la membrane. La liaison des ions chlorure induit un changement conformationnel dans la pompe, ce qui permet ├Ā une partie de la prot├®ine d'effectuer une phosphorylation. Cela se produit gr├óce ├Ā l'utilisation de l'ATP, o├╣ un groupement phosphate est transf├®r├® ├Ā la pompe. Le processus de phosphorylation favorise le transport des ions chlorure ├Ā travers la membrane, contre leur gradient ├®lectrochimique. Cela signifie que la pompe travaille activement pour accumuler les ions chlorure de part et d'autre de la membrane, cr├®ant ainsi un gradient de concentration. Apr├©s le transport des ions chlorure, la pompe reprend sa conformation initiale gr├óce ├Ā la d├®phosphorylation, qui est catalys├®e par une enzyme phosphatase associ├®e. Ce processus permet de pr├®parer la pompe pour de nouveaux cycles de liaison d'ions chlorure et de transport. Pompes
├Ā cations divalents.
Au repos, la pompe ├Ā cations divalents est li├®e ├Ā une mol├®cule d'ad├®nosine triphosphate (ATP), une source d'├®nergie cellulaire. L'ATP est hydrolys├®e pour fournir l'├®nergie n├®cessaire au fonctionnement de la pompe. Les ions calcium ou magn├®sium se lient ├Ā des sites sp├®cifiques sur la prot├®ine, g├®n├®ralement situ├®s du c├┤t├® cytoplasmique (intracellulaire) de la membrane. La liaison des ions cations divalents induit un changement conformationnel dans la pompe, ce qui permet ├Ā une partie de la prot├®ine d'effectuer une phosphorylation. Cela se produit gr├óce ├Ā l'utilisation de l'ATP, o├╣ un groupement phosphate est transf├®r├® ├Ā la pompe. Le processus de phosphorylation favorise le transport des ions cations divalents ├Ā travers la membrane, contre leur gradient ├®lectrochimique. Cela signifie que la pompe travaille activement pour accumuler les ions cations divalents de part et d'autre de la membrane, cr├®ant ainsi un gradient de concentration. Apr├©s le transport des ions cations divalents, la pompe reprend sa conformation initiale gr├óce ├Ā la d├®phosphorylation, qui est catalys├®e par une enzyme phosphatase associ├®e. Ce processus permet de pr├®parer la pompe pour de nouveaux cycles de liaison d'ions cations divalents et de transport. Autres
transports actifs par transporteur.
ŌĆó Transporteur de glucose. - Ce transporteur est responsable du mouvement du glucose contre son gradient de concentration ├Ā travers la membrane cellulaire, dans certains tissus, comme les cellules musculaires ou adipeuses, permettant ainsi l'absorption du glucose m├¬me lorsque sa concentration extracellulaire est faible.Cotransport. Le cotransport utilise comme source d'├®nergie le gradient ├®lectrochimique d'une substance (g├®n├®ralement un ion) cr├®├® par un transport actif ├Ā contre-gradient pour le transport d'une autre substance contre son gradient de concentration. On est ainsi conduit ├Ā distinguer un transport actif primaire (le transport actif ├Ā contre-gradient) et un transport secondaire : ŌĆó Transport actif primaire. - Dans le transport actif primaire, comme on l'a vu, l'├®nergie directe fournie par l'hydrolyse de l'ad├®nosine triphosphate (ATP) est utilis├®e pour d├®placer les substances ├Ā travers la membrane cellulaire, contre leur gradient de concentration. La substance est d├®plac├®e de la r├®gion de concentration plus basse vers celle de concentration plus ├®lev├®e.Il existe deux types principaux de cotransport, le cotransport symport et le cotransport antiport. Dans le premier, deux substances sont transport├®es dans la m├¬me direction ├Ā travers la membrane, tandis que dans le second, les substances sont transport├®es dans des directions oppos├®es : Le
cotransport symport.
ŌĆó Absorption de glucose et de sodium. - Dans les racines des plantes, il existe des cotransporteurs symport qui permettent l'absorption simultan├®e de glucose et de sodium (NaŌü║) ├Ā travers la membrane plasmique. Le glucose est transport├® dans la cellule en utilisant le gradient de sodium├®tabli par une pompe ├Ā sodium ATP-d├®pendante (transport actif primaire). Dans les cellules intestinales et r├®nales, o├╣ le glucose est co-transport├® avec le sodium ├Ā travers la membrane cellulaire. Le gradient de sodium existant ├Ā travers la membrane fournit l'├®nergie n├®cessaire pour transporter le glucose contre son propre gradient de concentration (transport actif secondaire). Cela permet d'absorber efficacement le glucose des aliments dans l'intestin ou de le r├®absorber dans les reins. ŌĆó Transport chlorure-bicarbonate. - Ce cotransport symport se produit ici dans les ├®rythrocytes (globules rouges) o├╣ le chlorure et le bicarbonate sont transport├®s dans le m├¬me sens ├Ā travers la membrane plasmique. Ce processus est important pour le transport du dioxyde de carbone dans le sang et pour maintenir l'├®quilibre acido-basique de l'organisme.Le cotransport antiport. Dans le cotransport antiport (= cotransport antiporteur) deux substances sont transport├®es simultan├®ment ├Ā travers une membrane cellulaire, mais dans des directions oppos├®es. Le cotransport antiport utilise l'├®nergie d'un gradient ├®lectrochimique pr├®existant pour entra├«ner le transport actif des substances contre leur gradient de concentration. Exemples : ŌĆó R├®gulation du pH cellulaire. - Dans les cellules v├®g├®tales, il existe des cotransporteurs antiport qui transportent simultan├®ment des ions HŌü║ (protons) hors de la cellule contre leur gradient de concentration (transport actif primaire), tout en transportant un autre ion, comme le potassium (KŌü║), dans la cellule (transport actif secondaire). ŌĆó ├ēchange Calcium-Sodium. - Ce cotransport antiport se produit dans les membranes des cellules musculaires cardiaques et lisses. Il utilise l'├®nergie du gradient de sodium pour extraire le calcium de la cellule. C'est un processus qui intervient dans la relaxation musculaire et la r├®gulation de la contraction. |
| . |
|
|
|||||||||||||||||||||||||||||||
|
