 |
Les neurones
ou cellules nerveuses sont, avec les cellules
gliales (qui sont d'abord des cellules de soutien), les cellules
dont est constitué le système nerveux.
Les neurones sont responsables de la transmission de l'information sous
forme de signaux électriques et chimiques et communiquent des informations
sur les sensations et qui produisent des mouvements en réponse à ces
stimuli, tout en induisant des processus de pensée
dans le cerveau. Une partie importante du fonctionnement
des neurones réside dans leur structure ou leur forme. La forme tridimensionnelle
de ces cellules rend possible l'immense nombre de connexions au sein du
système nerveux.
Chaque neurone est
composé d'un corps cellulaire (soma) contenant le noyau et divers organites,
et de prolongements : les dendrites en nombre variable et l'axone, qui
est unique unique. Les dendrites sont une série de prolongements rayonnants,
qui relient directement le neurone considéré aux neurones qui l'environnent;
permettant le passage des autres neurones, tandis que l'axone transmet
les signaux à d'autres neurones ou à des cellules cibles d'un quelconque
organe effecteur (muscle, en particulier) ou de
glandes.Les neurones communiquent entre eux par le biais de synapses,
des jonctions sp√©cialis√©es o√Ļ les signaux √©lectriques sont convertis
en signaux chimiques (neurotransmetteurs) pour être transmis à d'autres
neurones.
Les parties des neurones
La partie principale
d'un neurone est le corps cellulaire, également connu sous le nom de soma
(= corps). Le corps cellulaire contient le noyau et la plupart des principaux
organites. Mais ce qui rend les neurones spéciaux, c'est qu'ils
possèdent de nombreuses extensions ou prolongements de leurs membranes
cellulaires : l'axone et les dendrites. Cet axone unique peut se
ramifier à plusieurs reprises pour communiquer avec de nombreuses cellules
cibles. C'est l'axone qui qui transmet les signaux électriques à d'autres
neurones ou à des cellules cibles. Les dendrites, quant à elles, reçoivent
des informations d'autres neurones au niveau de zones de contact spécialisées
appelées synapses. Les dendrites sont généralement des prolongements
très ramifiés, fournissant des emplacements permettant à d'autres neurones
de communiquer avec le corps cellulaire. L'information circule ainsi à
travers un neurone depuis les dendrites, à travers le corps cellulaire
et jusqu'à l'axone. Cela confère au neurone une polarité, ce qui signifie
que les informations circulent dans cette seule direction.
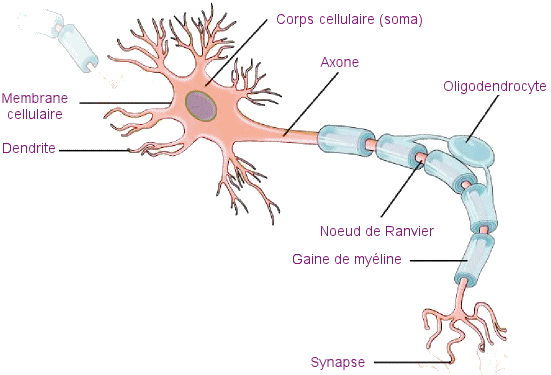
Schéma
des différentes parties d'un neurone typique (neurone multipolaire).
(Source
: Document Openstax).
Le corps cellulaire.
Le corps cellulaire
( = soma) est la partie centrale du neurone o√Ļ se trouvent la plupart
des organites cellulaires que l'on rencontrerait dans n'importe quel autre
type de cellule, et qui sont responsables du métabolisme cellulaire
et de la synthèse des protéines nécessaires au fonctionnement de la
cellule. Le corps cellulaire des neurones présente seulement quelques
particularit√©s structurales et fonctionnelles adapt√©es √† leur r√īle
dans la transmission et le traitement de l'information dans le système
nerveux le corps cellulaire des neurones contient un noyau volumineux et
bien développé et le réticulum endoplasmique rugueux est abondant. Ces
deux organites sont impliqués dans la production de protéines traduisent
notamment le fait que les neurones nécessitent une quantité considérable
de protéines, synthétisées rapidement, pour maintenir leur structure,
fonctionner correctement et communiquer avec d'autres neurones. On note
aussi que, dans le corps cellulaire des neurones, les mitochondries
sont particulièrement nombreuses (elles sont aussi présentes dans
les dendrites et l'axone). Ce qui s'explique par le fait que les neurones
ont une demande énergétique élevée en raison de leurs fonctions spécialisées,
notamment, ici encore, la synthèse des protéines, le maintien du potentiel
de repos membranaire, le transport axonal et la transmission synaptique.
Or, cette demande énergétique élevée nécessite une production continue
d'ATP, ce qui est assuré par les mitochondries. Enfin, on note le caractère
dynamique particulier du cytosquelette des neurones, qui est la clé de
leur morphologie, de leur polarisation, de leur fonction de transport intracellulaire
et de leur plasticité synaptique.
Les dendrites.
Les dendrites sont
des prolongements cytoplasmiques, généralement très ramifiés, du corps
cellulaire des neurones, fournissant des emplacements permettant à d'autres
neurones de communiquer avec le corps cellulaire au niveau de zones de
contact spécialisées appelées synapses. Cela donne au neurone une polarité,
ce qui signifie que les informations circulent dans cette seule direction.
Les dendrites sont couvertes de récepteurs
spécialisés qui détectent les neurotransmetteurs
libérés par les terminaisons axonales d'autres neurones au niveau des
synapses. Ces récepteurs convertissent les signaux chimiques en signaux
électriques qui modifient le potentiel de membrane du neurone. L'intégration
des signaux excitateurs et inhibiteurs provenant de différentes synapses
se produit au niveau du corps cellulaire, o√Ļ les signaux sont somm√©s
pour déterminer si un potentiel d'action sera généré dans l'axone du
neurone. Les modifications de la structure et de la fonction des dendrites
peuvent être impliquées dans la formation de souvenirs et dans l'adaptation
du cerveau à de nouvelles expériences.
L'axone.
L'axone est une
longue extension unique du neurone qui émerge du corps cellulaire et
se projette vers les cellules cibles (de muscles ou de glandes). Cet axone
unique peut se ramifier à plusieurs reprises pour communiquer avec de
nombreuses cellules cibles. C'est l'axone qui propage l'influx nerveux
qui est communiqué à une ou plusieurs cellules. Les axones peuvent être
enrobés dans une gaine de myéline, isolante
électriquement et protectrice mécaniquement. La présence ou l'absence
de myéline autour de l'axone dépend du type de neurone et de son emplacement
dans le système nerveux.
‚ÄĘ Neurones
non-myélinisés. - Certains neurones, en particulier ceux du système
nerveux autonome, du système nerveux entérique et du système nerveux
périphérique sensoriel, sont souvent non myélinisés ou partiellement
myélinisés. Par exemple, les fibres C du système nerveux périphérique,
qui transmettent les signaux de douleur et de température.
‚ÄĘ Neurones partiellement
myélinisés. - Certains neurones peuvent être partiellement myélinisés,
avec seulement une partie de leur axone entourée par une gaine de myéline.
Cela peut être observé dans des neurones sensoriels du système nerveux
périphérique.
‚ÄĘ Neurones my√©linis√©s.
- D'autres neurones, en particulier ceux du système nerveux central impliqués
dans la transmission rapide des signaux, sont généralement fortement
myélinisés. Par exemple, les axones des neurones moteurs dans la moelle
épinière et les faisceaux de projection dans le cerveau.
À l'extrémité de
l'axone se trouve la terminaison axonale, o√Ļ les signaux √©lectriques
sont convertis en signaux chimiques sous forme de neurotransmetteurs, et
o√Ļ se trouvent g√©n√©ralement plusieurs branches s'√©tendant vers la cellule
cible, dont chacune se termine par un élargissement appelé bulbe d'extrémité
synaptique.
Les faisceaux d'axones,
entourés d'un tissu conjonctif et, dans de nombreux cas, également entourés
de couches de myéline pour une transmission efficace des signaux nerveux
sont appelés nerfs dans le système nerveux périphérique.
Ils servent transmettre des signaux sensoriels du corps vers le cerveau
(nerfs sensitifs ou afférents) ou des signaux moteurs du cerveau vers
les muscles ou les glandes (nerfs moteurs ou efférents). Lorsque ces faisceaux
d'axones appartiennent au système nerveux central , on les désigne sous
le nom de tractus. Les tractus relient différentes régions du cerveau
et de la moelle épinière, permettant la communication entre ces régions
et facilitant la coordination des fonctions du système nerveux central.
Les synapses.
Les synapses sont
les jonctions sp√©cialis√©es o√Ļ les neurones communiquent entre eux par
la transmission les
signaux chimiques ou électriques ou chimiques. Entre la membrane présynaptique
d'un neurone ( = la membrane de la terminaison axonale) de la membrane
postsynaptique d'un autre neurone ( = la membrane dendritique ou la membrane
cellulaire d'un autre type de cellule, comme une cellule musculaire ou
une cellule glandulaire) se situe un espace étroit, la fente synaptique,
remplie d'un liquide extracellulaire riche en glycoprotéines
et en autres molécules qui contribuent à la communication entre les neurones
pré- et postsynaptiques.
Types
de synapses.
Si l'on considère
les moyens par lesquels les synapses transmettent l'information, on peut
distinguer les synapses chimiques, qui sont de loin les plus courants dans
le système nerveux, et qui impliquent des neurotransmetteurs, et les synapses
électriques dans lesquelles le signal est transmis électriquement.
‚ÄĘ Synapses
chimiques. - Dans les synapses chimiques, la transmission du signal
se fait par le biais de neurotransmetteurs qui diffusent à travers la
fente synaptique et se lient à des récepteurs spécifiques sur la membrane
de la cellule postsynaptique, provoquant ainsi des changements dans le
potentiel de membrane postsynaptique, qui peuvent être excitatoires (augmentant
la probabilité de génération d'un potentiel d'action) ou inhibiteurs
(réduisant cette probabilité).
‚ÄĘ Synapses √©lectriquess.
- Dans de telles synapses, les cellules nerveuses sont physiquement connectées
par des jonctions dites jonctions communicantes. Celles-ci permettent un
passage direct d'ions et de molécules entre les cellules, favorisant une
transmission électrique rapide du signal.
Si lon considère les
synapses en fonction des parties des neurones qu'elles mettent en relation,
on peut distinguer les trois types de synapses suivants :
‚ÄĘ Les
synapses axo-axonales se situent entre l'axone d'une cellule présynaptique
et l'axone d'une cellule postsynaptique. Elles sont impliquées dans la
modulation de la transmission de l'information et peuvent influencer l'excitabilité
des neurones.
‚ÄĘ Les synapses
axo-somatiques se produisent entre l'axone d'une cellule présynaptique
et le corps cellulaire (soma) d'une cellule postsynaptique. Ces synapses
jouent un r√īle dans la r√©gulation de l'activit√© neuronale et peuvent
moduler l'intégration des signaux entrants dans le neurone postsynaptique.
‚ÄĘ Les synapses
axo-dendritiques sont formées entre l'axone d'une cellule présynaptique
et les dendrites d'une cellule postsynaptique. Ces synapses sont les plus
courantes et sont impliquées dans le traitement de l'information neuronale
et la régulation de l'activité cérébrale.
La
plasticité synaptique.
On parle de plasticité
synaptique pour désigner la capacité des synapses situées sur les dendrites
de modifier leur sensibilité et leur efficacité dans la transmission
des signaux, en réponse à des activités neuronales répétées ou à
des stimuli extérieurs. Cela revient en particulier à modifier les connexions
entre neurones. La plasticité synaptique est l'une des deux expressions
de plasticité neuronale ( = neuroplasticité), l'autre étant la
neurogénèse.
La plasticité synaptique est nécessaire à l'adaptation du cerveau à
son environnement et pour le traitement de l'information. Il existe plusieurs
formes de plasticité synaptique, parmi lesquelles :
‚ÄĘ La
potentialisation à long terme (LTP) implique le renforcement des connexions
synaptiques entre les neurones, souvent en réponse à une stimulation
intense et répétée. La LTP est largement considérée comme un mécanisme
central dans l'apprentissage et la mémoire.
‚ÄĘ La d√©pression
à long terme (LTD), contrairement à la LTP, implique une diminution
de l'activité des connexions synaptiques. Elle peut être déclenchée
par différentes conditions, comme une stimulation synaptique faible ou
persistante.
‚ÄĘ La plasticit√©
homéostatique permet au cerveau de maintenir un équilibre ou une
stabilité relative dans l'activité neuronale. Il ajuste l'efficacité
ou l'activité des synapses pour compenser les changements dans l'activité
neuronale globale, assurant ainsi la stabilité de la fonction cérébrale.
Classifications des
neurones
Il existe de nombreux
types de neurones. Ils peuvent être classés selon de nombreux critères
différents. Une façon de les classer consiste à considérer le nombre
de prolongements attachés au corps cellulaire (classification morphologique).
Les neurones peuvent également être classés selon leur fonction (classification
fonctionnelle), ou encore selon les substances chimiques qu'ils utilisent
pour communiquer entre eux (classification fonctionnelle selon les neurotransmetteurs).
Quelques mots donc sur ces classifications des neurones, qui sont parmi
les plus courantes, et surles bases sur lesquelles elles reposent. Elles
ne sont pas mutuellement exclusives et peuvent parfois se chevaucher.
Classification
morphologique.
Les neurones présentent
une grande diversité de formes et de structures, et ils peuvent être
classés en différents types morphologiques en fonction de la forme de
leur corps cellulaire, de leurs dendrites et de leur axone.
Neurones
unipolaires.
Les neurones unipolaires
proprements dits ne se rencontrent que chez les animaux invertébrés,
et sont un type de neurones caractérisés par la présence d'un seul prolongement
qui émerge du corps cellulaire, et l'absence de dendrites. Ce prolongement
se divise ensuite en deux branches distinctes, créant ainsi une structure
en forme de T. Ces neurones, principalement localisés dans le système
nerveux périphérique (SNP), en particulier dans les ganglions nerveux
dorsaux
de la moelle √©pini√®re et dans les ganglions des nerfs cr√Ęniens, sont
surtout impliqués dans la transmission des informations provenant des
récepteurs sensoriels périphériques (récepteurs cutanés de la pression
et de la température, récepteurs proprioceptifs responsables de la sensation
de position du corps). Dans ces neurones, la branche périphérique (qui
se connecte aux récepteurs sensoriels) transmet les signaux sensoriels
vers le corps cellulaire situé dans les ganglions nerveux, tandis que
la branche centrale (qui se prolonge vers le système nerveux central)
transporte les signaux vers le syst√®me nerveux central, o√Ļ ils sont trait√©s
et interprétés.
Neurones
pseudo-unipolaires.
Les neurones pseudo-unipolaires,
qui existent chez les Vertébrés, et particulièrement les humains, sont
un type spécial de neurones présents dans les ganglions spinaux et les
ganglions des nerfs cr√Ęniens. Ils sont caract√©ris√©s par une structure
unique o√Ļ l'axone et la dendrite fusionnent pour former un seul prolongement
qui se divise ensuite en deux branches distinctes. Cette caractéristique
fait que les neurones pseudo-unipolaires ressemblent à des neurones unipolaires,
mais ils diffèrent en termes de structure fonctionnelle. Contrairement
aux neurones unipolaires typiques, o√Ļ une branche est axonale et l'autre
est dendritique, les neurones pseudo-unipolaires ont un seul prolongement
qui fonctionne comme un axone et une dendrite en même temps. À
une extrémité de l'axone se trouvent des dendrites et à l'autre extrémité,
l'axone forme des connexions synaptiques avec une cible. Ainsi les signaux
sensoriels captés par les récepteurs périphériques (ex. : récepteurs
cutanés pour le toucher, la pression, etc.) sont transmis directement
le long de ce prolongement unique vers le corps cellulaire situé dans
les ganglions. À partir du corps cellulaire dans les ganglions, l'information
sensorielle est relayée vers le système nerveux central (cerveau et moelle
épinière) via une branche centrale du prolongement.
Neurones
bipolaires.
Les neurones bipolaires
possèdent deux prolongements distincts qui émergent à des extrémités
opposées du corps cellulaire : une dendrite et un axone. Ces neurones
ont une structure en forme de T avec une dendrite et un axone émergeant
de c√īt√©s oppos√©s du corps cellulaire. Les neurones bipolaires ne sont
pas très courants. On les trouve principalement dans l'épithélium olfactif
et dans la rétine. Dans la rétine, les neurones bipolaires sont situés
entre les photor√©cepteurs (c√īnes et b√Ętonnets) et les cellules ganglionnaires.
Ils reçoivent des signaux des photorécepteurs via leurs dendrites et
les transmettent ensuite aux cellules ganglionnaires via leur axone. Dans
la muqueuse olfactive, les neurones bipolaires jouent un r√īle similaire
en relayant les informations olfactives captées par les récepteurs olfactifs
aux cellules du bulbe olfactif du cerveau.
Neurones
multipolaires.
Les neurones multipolaires
sont tous les neurones qui ne sont ni unipolaires ni bipolaires. Ils ont
un axone et au moins deux dendrites (généralement beaucoup plus). À
l'exception des cellules ganglionnaires sensorielles unipolaires et des
cellules bipolaires spécifiques mentionnées ci-dessus, tous les autres
neurones sont multipolaires. Ces neurones sont les plus courants dans le
système nerveux central (SNC). Leur structure est adaptée à leur fonction
principale qui est l'intégration et le traitement de l'information neuronale.
Les dendrites reçoivent des signaux provenant d'autres neurones, sous
forme d'impulsions électriques ou chimiques, et les transmettent vers
le corps cellulaire. Une fois que l'information est intégrée au niveau
du corps cellulaire, elle est ensuite transmise le long de l'axone vers
d'autres neurones ou vers des muscles ou des glandes, selon le type de
neurone et la fonction qu'il remplit. Il existe plusieurs sous-types de
neurones multipolaires, distingués en fonction de leur localisation, du
nombre de dendrites et de l'organisation de leurs prolongements axonaux.
Exemples :
‚ÄĘ Neurones
pyramidaux. - Ils sont ainsi nommés en raison de la forme pyramidale
de leur corps cellulaire. Les neurones pyramidaux sont abondants dans le
cortex cérébral et ont une grande dendrite apicale qui se projette vers
le haut, ainsi que de nombreuses dendrites plus petites émergeant de leur
corps cellulaire. Ils sont impliqués dans des fonctions cognitives complexes
telles que l'apprentissage, la m√©moire et le contr√īle moteur fin.
‚ÄĘ Neurones stellaires.
- Ils ont une forme étoilée avec quelques dendrites qui émergent du
corps cellulaire. Les neurones stellaires sont présents dans le cervelet
et sont impliqués dans la modulation des signaux transmis par d'autres
neurones.
‚ÄĘ Neurones de
Purkinje. - Ces neurones se trouvent dans la couche de Purkinje du
cervelet. Ils ont un grand corps cellulaire avec de nombreuses dendrites
ramifiées en forme d'éventail. Les neurones de Purkinje interviennent
dans la coordination des mouvements et l'apprentissage moteur.
‚ÄĘ Neurones granulaires.
- Présents dans le cortex cérébelleux,
ces neurones ont de petites tailles de corps cellulaire avec de nombreuses
dendrites courtes. Ils forment la couche granulaire du cortex cérébelleux
et sont impliqués dans le traitement et l'intégration des informations
sensorielles et motrices.
‚ÄĘ Neurones fusiformes.
- Ces neurones se trouvent dans différentes régions du cerveau, en particulier
dans le cortex cérébral et le système
limbique. Ils ont une forme allongée avec une dendrite principale
et des dendrites secondaires qui émergent de leur corps cellulaire. Les
neurones fusiformes sont impliqués dans notamment la perception sensorielle,
les émotions et la mémoire.
‚ÄĘ Neurones moteurs.
- Les neurones moteurs (motoneurones) autonomes et ceux des nerfs cr√Ęniens,
ainsi que la plupart des motoneurones de la corne antérieure de la moelle
√©pini√®re (qui sont responsables du contr√īle moteur volontaire) ont une
structure multipolaire.
‚ÄĘ Neurones anaxoniques.
- Ces neurones sont très petits. Observés microscope, à la résolution
standard utilisée en histologie (environ 400X à 1000X de grossissement),
ils ne présentent aucun prolongement identifiable spécifiquement comme
un axone. En fait, n'importe lequel de ces prolongement pourrait fonctionner
comme un axone en fonction des conditions du moment.
Classification fonctionnelle.
Globalement, les
neurones sont spécialisés dans différentes fonctions selon leur structure
et de leur localisation, mais de nombreux neurones peuvent remplir plusieurs
fonctions selon lecontexte et les signaux auxquels ils sont exposés.
Neurones
sensoriels.
Les neurones sensoriels
( = neurones afférents), sont une classe de neurones responsables de la
transmission des signaux sensoriels provenant des organes sensoriels vers
le système nerveux central. Ils sont spécialisés dans la détection
des stimuli externes et internes, tels que la lumière, le son, la pression,
la température, la douleur, les substances chimiques, etc. Une fois qu'un
neurone sensoriel est activé par un stimulus, il génère un signal électrique
(potentiel d'action), qui est ensuite transmis le long de l'axone vers
le système nerveux central pour traitement. Les neurones sensoriels sont
extrêmement diversifiés en fonction de leur fonction sensorielle spécifique.
Par exemple, les neurones sensoriels de la rétine sont spécialisés dans
la détection de la lumière et la vision, tandis que les neurones sensoriels
du système olfactif sont spécialisés dans la détection des odeurs.
Neurones
moteurs.
Les neurones moteurs
( = neurones efférents = motoneurones) sont un type de neurones spécialisés
dans la transmission des signaux sous forme d'influx nerveux électriques
le long de leurs axones, jusqu'√† leurs terminaisons synaptiques o√Ļ ils
établissent des connexions avec les muscles ou les glandes cibles. Là,
ils déclenchent une réponse appropriée, provoquant la contraction musculaire
ou la sécrétion glandulaire nécessaire pour répondre aux stimuli ou
aux commandes reçus du système nerveux central. Les neurones moteurs
somatiques contr√īlent les muscles squelettiques volontaires. Les neurone
moteurs autonomes contr√īlent les fonctions involontaires. De mani√®re
plus fin, on distingue généralement plusieurs catégories de neurones
moteurs :
‚ÄĘ Les
motoneurones supérieurs sont un groupe de neurones moteurs situés
dans le cerveau, en particulier dans le cortex moteur primaire et d'autres
r√©gions corticales associ√©es au contr√īle moteur. Ils jouent un r√īle
essentiel dans l'initiation, la modulation et la coordination des mouvements
volontaires du corps.
+ Les
neuromoteurs corticaux ( = neurones de Betz) sont des cellules nerveuses
situées dans le cortex moteur primaire du cerveau ( = aire motrice primaire
= gyrus précentral). Ils sont responsables
de l'initiation et de la planification des mouvements volontaires. Ces
neurones sont caractérisés par leur grande taille et leur localisation
dans la couche V du cortex moteur primaire. Ils envoient des axones qui
descendent dans la moelle épinière et font partie du tractus corticospinal
(ou tractus pyramidal). Les neurones de Betz sont particulièrement importants
dans le contr√īle des mouvements fins et pr√©cis des muscles distaux, tels
que ceux des mains et des doigts.
+ Les
motoneurones des nerfs cr√Ęniens sont situ√©s dans les noyaux
moteurs des nerfs cr√Ęniens du tronc c√©r√©bral. Ces neurones sont responsables
de l'innervation des muscles moteurs des yeux, de la face, de la langue
et des muscles de la mastication, entre autres. Ils transmettent les impulsions
nerveuses du cerveau vers les muscles pour contr√īler les mouvements et
les fonctions des diff√©rentes structures cr√Ęniennes.
+
On peut encore mentionner d'autres catégoories motoneurones supérieurs,
qui contribuent tous de manière significative à la régulation et au
contr√īle des mouvements volontaires, des r√©flexes et de la posture du
corps en coordonnant les signaux provenant du cortex moteur, du cervelet,
du système vestibulaire et d'autres régions du cerveau-:
les motoneurones vestibulospinaux, situés dans les noyaux vestibulaires
du tronc cérébral quisont responsables de la régulation des réflexes
vestibulo-spinaux et contr√īlent l'√©quilibre et la posture en r√©ponse
aux mouvements de la tête; les motoneurones rubrospinaux,
situés dans le noyau rouge du mésencéphale
et qui envoient des projections descendantes vers la moelle épinière
pour contribuer à la régulation des mouvements des membres et de la posture;
les motoneurones réticulospinaux, situés dans le système réticulaire
du tronc cérébral et qui contribuent à la modulation des réflexes spinaux,
à la régulation de la posture et à la coordination des mouvements; les
motoneurones
tectospinaux, sont situés dans le colliculus supérieur du mésencéphale,
qui sont impliqués dans les réponses aux stimuli visuels, en particulier
les mouvements oculaires associés à l'orientation du regard.
‚ÄĘ Motoneurones
de la corne antérieure de la moelle épinière ( = neurones moteurs
inférieurs = neurones moteurs spinaux). - Ces motoneurones envoient
leurs axones directement aux muscles squelettiques striés, provoquant
leur contraction. Ils jouent un r√īle essentiel dans la g√©n√©ration
du mouvement volontaire.
+ Les
motoneurones Alpha sont les principaux neurones moteurs inféreurs.
Ils innervent des fibres musculaires extrafusales, qui sont responsables
de la contraction musculaire et du mouvement volontaire. Les motoneurones
alpha sont connectés aux fibres musculaires ordinaires (extrafusales)
et déclenchent leur contraction en réponse à des signaux provenant du
cerveau ou de la moelle épinière.
+ Les motoneurones
Gamma sont responsables de l'innervation des fibres musculaires intrafusales,
qui sont des fibres musculaires spécialisées présentes dans les fuseaux
neuromusculaires, des organes sensibles à l'étirement musculaire. Les
motoneurones gamma régulent la sensibilité des fuseaux neuromusculaires
en maintenant une tension appropriée dans les fibres intrafusales, ce
qui contribue à la régulation de la posture et du tonus musculaire.
+ Les motoneurones
B√™ta sont √©galement impliqu√©s dans le contr√īle moteur, mais leur
fonction spécifique peut varier selon le contexte. En général, ils innervent
les fibres musculaires moins rapides que les motoneurones alpha et peuvent
√™tre impliqu√©s dans le contr√īle plus fin des mouvements ou dans le recrutement
de fibres musculaires spécifiques.
‚ÄĘ Les neurones
moteurs autonomes ( = neurones moteurs viscéraux = neurones
moteurs des ganglions autonomes), contrairement aux neurones des types
pr√©c√©dents (neurones moteurs somatiques) qui contr√īlent les mouvements
volontaires des muscles squelettiques, les neurones moteurs autonomes contr√īlent
les muscles lisses, les muscles cardiaques et les glandes. Les motoneurones
autonomes régulent ainsi des fonctions involontaires du corps. Exemples
:
+ Dans
le système
cardiovasculaire, les neurones moteurs
autonomes contr√īlent la fr√©quence cardiaque, la force de contraction
du muscle cardiaque et la distribution du sang dans les vaisseaux sanguins.
+ Dans l'appareil
respiratoire, ils régulent le diamètre des voies respiratoires
et influencent la fréquence respiratoire.
+ Dans l'appareil
digestif, ils contr√īlent la motilit√© gastro-intestinale, la s√©cr√©tion
de sucs digestifs et la vasodilatation dans le tractus gastro-intestinal.
+ Dans l'appareil
urinaire, ils influent sur la contraction de la vessie et des muscles
lisses des voies urinaires.
+ Dans le système
endocrinien, ils modulent la sécrétion d'hormones
par les glandes endocrines telles que l'hypophyse,
la glande thyro√Įde et les glandes surr√©nales.
Les neurones moteurs
font partie du système nerveux autonome (SNA).
Le système nerveux
autonome étant partagé en deux subdivisions principales (le système
nerveux sympathique et le système nerveux parasympathique), ont peut donc
également subdiviser les neurones moteurs autonomes en deux groupes principaux,
qui rendent comptent de leurs r√īles antagonistes et compl√©mentaires,
contribuant par là au maintien de l'homéostasie
interne et la régulation des fonctions automatiques du corps :
+ Les
neurones du système nerveux sympathique sont impliqués dans la mobilisation
des ressources corporelles en réponse au stress, à l'excitation ou à
des situations d'urgence. Ces neurones se projettent à partir de la moelle
épinière thoracique et lombaire. Les neurotransmetteurs principaux libérés
par
les neurones sympathiques sont la noradrénaline et, dans une moindre mesure,
l'adrénaline (également connue sous le nom d'épinéphrine). Les actions
typiques du système sympathique incluent l'augmentation de la fréquence
cardiaque, la dilatation des pupilles, la libération de glucose dans le
sang, la vasoconstriction dans la peau et la relaxation des muscles lisses
dans les voies respiratoires.
+ Les neurones
du système nerveux parasympathique sont impliqués dans la conservation
et la restauration de l'énergie corporelle, favorisant des états de repos,
de digestion et de récupération. Ces neurones se projettent à partir
de la moelle épinière sacrée et du tronc cérébral (notamment le nerf
vague). Le neurotransmetteur principal libéré par les neurones
parasympathiques est l'acétylcholine. Les actions typiques du système
parasympathique incluent la diminution de la fréquence cardiaque, la contraction
des pupilles, la stimulation de la digestion et la vasodilatation dans
la peau.
Les
neurones intermédiaires.
Les neurones intermédiaires
( = interneurones = neurones d'association) sont un type de neurones présents
principalement dans le système nerveux central et qui agissent comme des
intermédiaires dans le traitement de l'information neuronale. Parmi leurs
fonctions, on mentionnera :
‚ÄĘ L'int√©gration
sensorielle. - Les neurones intermédiaires reçoivent des signaux
provenant des neurones sensoriels et les intègrent pour générer des
réponses appropriées. Par exemple, dans le réflexe
rotulien, les neurones intermédiaires de la moelle épinière intègrent
les signaux provenant des neurones sensoriels et déclenchent une réponse
de contraction du muscle quadriceps.
‚ÄĘ La modulation
de l'activité neuronale. - Les neurones intermédiaires peuvent modifier
l'activité des autres neurones en régulant l'intensité et la fréquence
des signaux transmis. Ils agissent comme des intermédiaires dans la transmission
de l'information entre les neurones sensoriels et moteurs, ainsi que dans
la régulation des réponses neuronales.
‚ÄĘ La coordination
des mouvements. - Les neurones intermédiaires participent à la coordination
des mouvements en intégrant les signaux provenant de différentes régions
du système nerveux central et en synchronisant l'activité des muscles
pour produire des mouvements fluides et coordonnés.
Les
neurones miroirs.
Les neurones miroirs
sont un type particulier de neurones qui s'activent à la fois quand on
exécute une action et quand on observe quelqu'un d'autre faire cette même
action. Autrement dit, le cerveau ‚Äúsimule‚ÄĚ int√©rieurement ce qu'il
voit, comme s'il le faisait lui-même. Ils ont été identifiés dans les
années 1990 par l'équipe du neuroscientifique
Giacomo Rizzolatti, en étudiant le cerveau de singes. Les chercheurs ont
remarqué que certains neurones s'activaient aussi bien quand le singe
attrapait un objet que lorsqu'il voyait un autre individu effectuer ce
geste.
Chez l'humain, ces
neurones sont impliqués dans plusieurs fonctions clés. Ils jouent un
r√īle dans l'apprentissage par imitation, ce qui explique pourquoi on peut
apprendre des gestes simplement en observant (par exemple dans le sport
ou les arts). Ils participent aussi à la compréhension des intentions
des autres, en ‚Äúreproduisant‚ÄĚ mentalement leurs actions. Enfin, ils
sont souvent associés à l'empathie, car ils contribuent à ressentir
ou à comprendre les émotions d'autrui en activant des circuits similaires
dans notre propre cerveau.
Cela dit, leur r√īle
exact reste débattu. Certains chercheurs leur attribuent une importance
centrale dans des capacités complexes comme le langage
ou la cognition sociale, tandis que d'autres
estiment que leur fonction est plus limitée et qu'ils ne suffisent pas,
à eux seuls, à expliquer ces phénomènes.
La classification
fonctionnelle basée sur les neurotransmetteurs.
La classification
fonctionnelle basée sur les neurotransmetteurs catégorise les neurones
en fonction des neurotransmetteurs qu'ils libèrent comme principaux moyens
de communication avec d'autres neurones. Mais on notera que certains neurones
possèdent les enzymes nécessaires pour synthétiser
différents neurotransmetteurs dans différentes parties de leur structure,
ce qui ajoute une complexité au système nerveux en permettant une modulation
fine et précise de la
signalisation
neuronale en réponse à différentes conditions physiologiques et environnementales.
Ces neurones sont appelés neurones multifonctionnels ou neurones
co-transmetteurs.
 La
classification fonctionnelle basée sur les neurotransmetteurs catégorise
les neurones en fonction des neurotransmetteurs qu'ils libèrent comme
principaux moyens de communication avec d'autres neurones. Ainsi, peut-on
distinguer les
neurotransmetteurs excitateurs déclenchent ou amplifient
les signaux électriques dans les neurones postsynaptiques (exemples :
glutamate, acétylcholine); les
neurotransmetteurs inhibiteurs réduisent
ou inhibent l'activité électrique des neurones postsynaptiques (ex. :
GABA, glycine); les neurotransmetteurs modulateurs peuvent augmenter
ou réduire l'effet des neurotransmetteurs excitateurs ou inhibiteurs,
modulant ainsi l'activité d'autres neurotransmetteurs (ex. : dopamine,
sérotonine,
noradrénaline). La
classification fonctionnelle basée sur les neurotransmetteurs catégorise
les neurones en fonction des neurotransmetteurs qu'ils libèrent comme
principaux moyens de communication avec d'autres neurones. Ainsi, peut-on
distinguer les
neurotransmetteurs excitateurs déclenchent ou amplifient
les signaux électriques dans les neurones postsynaptiques (exemples :
glutamate, acétylcholine); les
neurotransmetteurs inhibiteurs réduisent
ou inhibent l'activité électrique des neurones postsynaptiques (ex. :
GABA, glycine); les neurotransmetteurs modulateurs peuvent augmenter
ou réduire l'effet des neurotransmetteurs excitateurs ou inhibiteurs,
modulant ainsi l'activité d'autres neurotransmetteurs (ex. : dopamine,
sérotonine,
noradrénaline).
Quelques-uns des principaux
neurotransmetteurs et les systèmes de neurones impliqués dans leur production.
:
-
| ‚ÄĘ
Acétylcholine,
(ACh). - Principalement produite par les neurones cholinergiques dans le
système nerveux central et le système nerveux périphérique. Impliquée
dans la régulation de la mémoire, de l'attention, de la fonction musculaire
et du sommeil.
‚ÄĘ Dopamine.
- Produite par les neurones dopaminergiques dans le système nerveux central.
Impliquée dans la régulation de l'humeur, de la motivation, de la récompense,
du mouvement et de la cognition.
‚ÄĘ Noradr√©naline
(ou norépinéphrine). - Produite par les neurones noradrénergiques dans
le système nerveux central et le système nerveux périphérique. Impliquée
dans la régulation de l'attention, de l'humeur, du stress et de la vigilance.
‚ÄĘ Glycine.
- Principalement produit par les neurones glycinergiques de la moelle épinière
et du tronc cérébral. Principal neurotransmetteur inhibiteur dans la
moelle épinière, intervenant dans la régulation de la transmission des
signaux sensoriels et moteurs. |
‚ÄĘ
GABA
(acide gamma-aminobutyrique). - Principalement produit par les neurones
GABAergiques dans le système nerveux central. Il s'agit du principal neurotransmetteur
inhibiteur dans le cerveau. Il est impliqué dans la régulation de l'excitabilité
neuronale.
‚ÄĘ Glutamate.
- Principalement produit par les neurones glutamatergiques dans le système
nerveux central. C'est le principal neurotransmetteur excitateur dans le
cerveau, impliqué dans de nombreuses fonctions cérébrales (apprentissage,
la mémoire, etc.).
‚ÄĘ S√©rotonine.
- Produite par les neurones sérotoninergiques dans le système nerveux
central. Impliquée dans la régulation de l'humeur, du sommeil, de l'appétit,
de la douleur et de la fonction digestive.
‚ÄĘ Histamine.
- Principalement produite par les neurones histaminergiques dans le système
nerveux central. Impliquée dans la régulation de la vigilance, du sommeil
et de la réponse immunitaire. |
Neurones
sécrétant de la dopamine.
Les neurones sécrétant
de la dopamine comme neurotransmetteur
principal sont impliqués dans des voies
neurologiques importantes, notamment dans le système de récompense
du cerveau. La dopamine intervient √©galement dans le contr√īle des mouvements,
la motivation et la cognition. Des dysfonctionnements des neurones dopaminergiques
sont associés à des troubles tels que la maladie de Parkinson et la schizophrénie.
Neurones
sécrétant de la sérotonine.
Les neurones sécrétant
de la sérotonine comme neurotransmetteur
principal interviennent dans la modulation de l'humeur et sont souvent
ciblés dans le traitement des troubles de l'humeur tels que la dépre.
Ils sont également impliqués dans la régulation du cycle veille-sommeil
et de l'appétit.
Neurones
sécrétant de l'acétylcholine.
Les
neurones qui sécrètent l'acétylcholine
comme neurotransmetteur principal sont répartis dans tout le système
nerveux, o√Ļ ils participent notamment au contr√īle des muscles squelettiques,
à la modulation de l'activité cérébrale et à la transmission des signaux
dans le système nerveux autonome. Ils sont également impliqués dans
les processus de mémoire et d'apprentissage.
Neurones
sécrétant du GABA.
Les neurones sécrétant
du GABA (acide gamma-aminobutyrique) comme
neurotransmetteur principal (= neurones GABAergiques) sont des neurones
inhibiteurs. Ils sont abondants dans le cerveau et la moelle épinière,
o√Ļ ils modulent l'activit√© des neurones excitateurs et contribuent √†
maintenir l'homéostasie neuronale. Des dysfonctionnements des neurones
GABAergiques sont associés à des troubles neurologiques tels que l'épilepsie
et les troubles anxieux.
Communications entre
neurones
La communication entre
les neurones se fait par le biais de signaux électriques et chimiques,
qui sont un changement brusque de la polarité électrique de la membrane
cellulaire. Un tel signal, appelé influx nerveux ou potentiel
d'action, est g√©n√©r√© au niveau du corps cellulaire ou du c√īne
d'implantation de l'axone et se progage le long de l'axone. Lorsqu'il en
atteint l'extrémité, il déclenche la libération de neurotransmetteurs
dans l'espace synaptique. Les neurotransmetteurs se lient ensuite
aux récepteurs présents sur la membrane postsynaptique du neurone récepteur.
Cette liaison neurotransmetteur-récepteur déclenche des modifications
électriques dans le neurone postsynaptique, modulant son potentiel de
membrane. Les signaux électriques postsynaptiques générés dans le neurone
récepteur sont intégrés avec d'autres signaux provenant d'autres synapses.
Si la somme des signaux excitateurs et inhibiteurs atteint un seuil critique,
cela peut déclencher la génération d'un potentiel d'action dans le neurone
postsynaptique, qui se se propagera à son tour le long de son axone. Ces
étapes de la communication neuronale se répètent de manière coordonnée,
permettant le traitement de l'information et la régulation des fonctions
physiologiques et comportementales.
Génération du
potentiel d'action.
L'influx nerveux
appelé à parcourir l'axone est déclenché par l'appartition d'un potentiel
d'action. Avant la génération de ce potentiel, la membrane
cellulaire est polarisée, avec une charge négative à l'intérieur
de la cellule par rapport √† l'ext√©rieur. Cela est d√Ľ √† la distribution
asymétrique des ions à travers la membrane, notamment la concentration
plus élevée de sodium (Na+) et de
chlore
(Cl-) à l'extérieur et de
potassium
(K+) à l'intérieur.
Le potentiel d'action
appara√ģt g√©n√©ralement √† la suite d'une stimulation externe ou interne.
Cette stimulation peut être chimique, électrique ou mécanique. Lorsqu'un
elle est suffisammente forte, elle provoquera une dépolarisation de la
membrane au niveau d'une région spécifique, souvent près de l'endroit
o√Ļ se trouve le corps cellulaire (soma) ou le d√©but de l'axone. Cette
dépolarisation peut résulter de l'ouverture de canaux
ioniques spécifiques, généralement des canaux sodium voltage-dépendants.
Si la dépolarisation atteint un seuil critique, généralement autour
de -55 millivolts (mV) par rapport au potentiel de repos, cela déclenche
l'ouverture massive et rapide d'autres canaux sodium voltage-dépendants
et à un grand nombre d'ions sodium (Na+)
entrent dans la cellule, ce qui entra√ģne une
inversion soudaine du potentiel électrique de la membrane. Le potentiel
d'action peut commencer à se propager.
Propagation du
potentiel d'action..
Une fois déclenché,
le potentiel d'action se propage de manière uniforme et auto-entretenue
le long de l'axone du neurone, sans perte de force, gr√Ęce √† un processus
appelé régénération, qui découle de l'ouverture séquentielle des
canaux sodiques voltage-dépendants le long de l'axone,
provoquant ainsi la dépolarisation de la membrane à chaque segment de
l'axone. La membrane doit rétablir ensuite son état de repos initial.
Les canaux sodiques voltage-dépendants se ferment et les canaux potassiques
voltage-d√©pendants s'ouvrent. Cela entra√ģne une sortie d'ions potassium
(K+) hors de la cellule, rétablissant
la polarisation de la membrane, c'est-à-dire son état de repos initial.
Elle est prête pour le prochain potentiel d'action.
Après le passage
du potentiel d'action, les segments de l'axone déjà dépolarisés deviennent
temporairement inexcitables. Une période réfractaire, qui garantit que
les signaux électriques se propagent de manière unidirectionnelle le
long de l'axone et qui contribue à la régulation de la fréquence des
impulsions neuronales.
Libération des
neurotransmetteurs.
Lorsque le potentiel
d'action atteint la terminaison axonique du neurone présynaptique, il
y déclenche l'ouverture des canaux calciques voltage-dépendants. L'entrée
de calcium dans la terminaison axonique provoque la fusion des vésicules
synaptiques contenant les neurotransmetteurs avec la membrane cellulaire,
ce qui libère les neurotransmetteurs dans l'espace synaptique. Ce processus
est appelé exocytose.
Un
neurone peut stocker différents neurotransmetteurs dans différentes
vésicules synaptiques, lui permettant de les libérer sélectivement en
fonction des besoins.Cette libération peut être régulée de manière
différentielle en fonction des signaux pré-synaptiques et des conditions
environnementales. Ainsi, un neurone peut libérer différents neurotransmetteurs
en réponse à différents stimuli.
Les neurotransmetteurs
libérés se diffusent à travers l'espace synaptique, entre la terminaison
axonique du neurone présynaptique et les dendrites ou le corps cellulaire
du neurone postsynaptique, et atteignent les récepteurs situés sur la
membrane postsynaptique. Ces récepteurs sont des protéines
spécifiques qui reconnaissent et se lient à un neurotransmetteur spécifique
ou un groupe de neurotransmetteurs ayant des structures chimiques similaires.
La structure chimique du neurotransmetteur est complémentaire à celle
du site de liaison du récepteur. Ils possèdent des formes spécifiques
qui leur permettent de se fixer de manière sélective aux récepteurs
correspondants, tout comme une clé s'insère dans une serrure. Une fois
que les neurotransmetteurs auront rempli leur r√īle, ils seront soit recycl√©s
par la cellule présynaptique pour une utilisation future, soit dégradés
par des enzymes présentes dans la fente synaptique.
Lorsque les neurotransmetteurs
se lient aux récepteurs postsynaptiques plusieurs effets peuvent s'observer.
Certains neurotransmetteurs, en se liant à leurs récepteurs postsynaptiques,
activent des voies de signalisation
intracellulaire qui peuvent déclencher des réponses cellulaires complexes,
telles que la modulation de l'activité des enzymes, la régulation de
l'expression génique ou la modification de la structure synaptique. La
stimulation répétée ou prolongée de certaines synapses peut aussi induire
des changements à long terme dans la plasticité synaptique. Les neurotransmetteurs
peuvent, enfin, moduler la conductance ionique postsynaptique en régulant
l'ouverture et la fermeture des canaux ioniques spécifiques, tels que
les canaux sodiques, potassiques, calciques ou chloriques. Cela induit
des changements dans la perméabilité de la membrane aux ions, affectant
ainsi le potentiel de membrane et la transmission du signal.
Certains
neurotransmetteurs, comme le glutamate, induisent la dépolarisation de
la membrane postsynaptique en ouvrant des canaux ioniques perméables aux
ions sodium (Na+) ou calcium (Ca2+).
La dépolarisation résulte en une augmentation du potentiel de membrane
postsynaptique, rapprochant ainsi le neurone post-synaptique du seuil d'excitation
nécessaire pour générer un potentiel d'action. D'autres neurotransmetteurs,
tels que le GABA (acide gamma-aminobutyrique) et la glycine, induisent
l'hyperpolarisation de la membrane postsynaptique en ouvrant des canaux
ioniques perméables aux ions chlorure
(Cl-) ou potassium
(K+). L'hyperpolarisation résulte en une
diminution du potentiel de membrane postsynaptique, éloignant ainsi le
neurone post-synaptique du seuil d'excitation.
Génération des
potentiels gradués.
Les changements
dans la perméabilité ionique de la membrane postsynaptique peuvent à
leur tour conduire à des modifications du potentiel de membrane au niveau
des dendrites en fonction du type de neurotransmetteur et des récepteurs
impliqués.On parle alors de potentiel gradué.
C'est un analogue du potentiel d'action, mais il est localisé, et son
déclenchement n'est défini par aucun seuil. La magnitude du potentiel
gradué dépend de l'intensité du stimulus et diminue rapidement avec
la distance du site de stimulation. Ces changements de tension électrique
sont souvent de faible amplitude et ne se propagent pas très loin le long
de la membrane.
Si le potentiel gradué
atteint un seuil critique, généralement autour de -55 mV à -50 mV dans
les neurones, il déclenche l'ouverture des canaux ioniques voltage-dépendants,
principalement des canaux sodium voltage-dépendants, dans la région de
l'axone du neurone. L'entrée massive d'ions sodium provoque une rapide
dépolarisation de la membrane, conduisant à un potentiel d'action, qui
se propagera le long de l'axone postsynaptique, poursuivant ainsi la transmission
du signal dans le système nerveux comme
on l'a vu précédemment.
Sans le cas o√Ļ le
potentiel gradué n'atteindrait pas le seuil nécessaire à la génération
d'un potentiel d'action, ce d√©clenchement reste possible gr√Ęce √† l'int√©gration
des signaux portés par plusieurs potentiels gradués. Le seuil d'excitation
nécessaire sera alors déterminé par la somme des potentiels membranaires
provenant de toutes les synapses excitatrices et inhibitrices. (La sommation
synaptique consiste à additionner les potentiels de membrane générés).
Il existe deux types principaux de sommation synaptique, la sommation temporelle
et la sommation spatiale :
‚ÄĘ Dans
la sommation temporelle, les potentiels de membrane produits par des
impulsions successives à partir de la même synapse ou de synapses voisines
sont additionnés dans le temps. Si plusieurs potentiels de membrane sont
générés suffisamment rapidement les uns après les autres, ils peuvent
se sommer pour dépasser le seuil d'excitation et déclencher un potentiel
d'action.
‚ÄĘ Dans la sommation
spatiale, les potentiels de membrane produits simultanément par différentes
synapses situées sur différentes régions du neurone postsynaptique sont
additionnés spatialement. Si plusieurs synapses excitées simultanément
contribuent suffisamment pour dépasser le seuil d'excitation, cela peut,
ici encore, entra√ģner la g√©n√©ration d'un potentiel d'action.
 |
Jean-Ga√ęl
Barbara, La naissance du neurone. Constitution d'un objet scientifique
au XXe siècle, Vrin, 2010.2711622614 |
|
|
