| . |
|
||||||
|
|
| . |
|
||||||
| Biologie > GĂŠnĂŠtique |
| L'ARN |
| L'acide
ribonuclĂŠique, ou ARN, est une molĂŠcule
d'acide nuclĂŠique
principalement impliquÊe dans le processus de synthèse des
protĂŠines
sous la direction de l'ADN. L'ARN est gĂŠnĂŠralement un simple brin et
est composĂŠ de ribonuclĂŠotides qui sont liĂŠs par des liaisons phosphodiester.
Un ribonuclĂŠotide dans la chaĂŽne d'ARN contient du ribose (sucre pentose),
l'une des quatre bases
azotĂŠes (A, U, G et C) et le groupe phosphate.
Il existe cinq principaux types d'ARN : l'ARN messager (ARNm), l'ARN ribosomal (ARNr), l'ARN de transfert (ARNt) et le microARN (miARN). L'ARN
messager.
La sÊquence de base de l'ARN est complÊmentaire de la sÊquence codante de l'ADN à partir de laquelle elle a ÊtÊ copiÊe. Cependant, dans l'ARN, la base T est absente et U est prÊsente à la place. Si le brin d'ADN a une sÊquence AATTGCGC, la sÊquence complÊmentaire de l'ARN est UUAACGCG. Dans le cytoplasme, l'ARNm interagit avec les ribosomes (structures à fonction catalytique, composÊes d'ARN et de protÊines) et d'autres machines cellulaires. L'ARNm est lu selon des sÊquences de trois bases appelÊes codons. Chaque codon code un seul acide aminÊ. De cette façon, l'ARNm est lu et le produit protÊique est fabriquÊ. L'ARN
ribosomal.
L'ARN
de transfert.
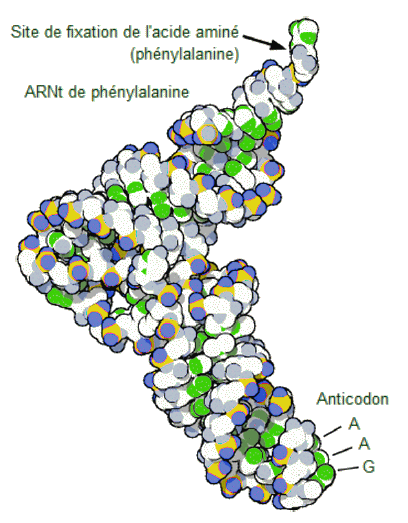
Les
microARN.
Les microARNs sont produits à partir de sÊquences spÊcifiques de l'ADN, transcrites sous forme de prÊcurseurs, appelÊs pri-miARNs, qui sont ensuite transformÊs en prÊ-miARNs puis en microARNs matures. Une fois matures, les microARNs s'associent à un complexe appelÊ RISC (RNA-induced silencing complex). Ils se lient ensuite de manière spÊcifique à des rÊgions complÊmentaires de l'ARN messager cible. Si cette complÊmentaritÊ est parfaite, cela peut entraÎner la dÊgradation de l'ARNm cible. Si la complÊmentaritÊ est imparfaite, ils inhibent la traduction de l'ARNm sans le dÊgrader, ce qui empêche la production de la protÊine. Les microARNs rÊgulent une grande variÊtÊ de processus biologiques. Ils interviennent dans la rÊgulation des gènes impliquÊs dans la croissance et le dÊveloppement. Ils jouent un rôle central dans la rÊgulation de la division cellulaire et de la mort cellulaire programmÊe (apoptose). Une dÊrÊgulation des microARNs peut entraÎner des maladies, notamment le cancer, oÚ certains microARNs peuvent agir comme des oncogènes (favorisant la croissance tumorale) ou comme des suppresseurs de tumeurs. Gènes et synthèse des protÊinesSelon l'hypothèse proposÊe par Crick dès 1958, il existe une relation entre l'ordre linÊaire des nuclÊotides dans l'ADN et l'ordre linÊaire des acides aminÊs dans les polypeptides (protÊines) à la synthèse desquelles il commande via l'ARN messager.On donne le nom de code gÊnÊtique à l'ensemble des règles qui dÊfinissent la manière dont l'information nÊcessaire à la synthèse des protÊines est transmise par l'ADN, et partant aux règles qui dÊfinissent la traduction des sÊquences nuclÊotidiques en sÊquences d'acides aminÊs dans une protÊine. Ce code est un "langage" qui repose sur plusieurs "alphabets" : l'alphabet ADN (A, T, C, G), l'alphabet ARN (A, U, C, G), et l'alphabet polypeptidique (20 acides aminÊs). Les "mots" sont composÊs de trois nuclÊotides. Les triplets de l'ARNm sont appelÊs codons et ceux de l'ARNt sont appelÊs anticodons. Chaque codon dÊtermine la nature de l'acide aminÊ qui sera synthÊtisÊ. Le code gÊnÊtique est dit redondant ou dÊgÊnÊrÊ, car les 4³ = 64 codons possibles dans l'ARNm ne spÊcifient que 20 acides aminÊs, un acide aminÊ donnÊ peut être codÊ par plus d'un triplet, et il y a en outre trois codons dits codons non-sens ou codons-stop, qui servent seulement à signaler la fin d'une chaÎne polypeptidique. Presque toutes les
espèces vivantes de la planète utilisent le même code gÊnÊtique.
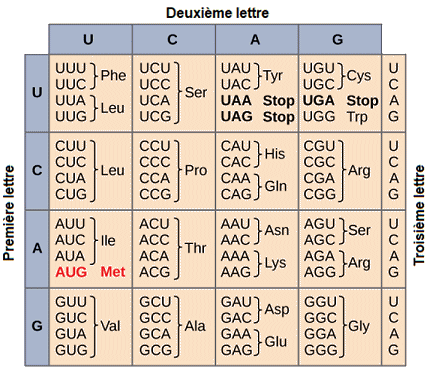
Si l'on poursuit la mÊtaphore grammaticale, on dira que le code gÊnÊtique mène à la construction de "phrases" : ce sont les gènes. Les gènes sont de petits segments d'ADN qui contiennent une sÊquence dÊfinie de nuclÊotides contenant toute l'information nÊcessaire pour fabriquer l'ARNm par un processus nommÊ transcription. L'ARNm est ensuite utilisÊ pour synthÊtiser les protÊines par le processus de traduction. La transcription.
Transcription
eucaryote.
Traitement
de l'ARN chez les eucaryotes.
Seuls les ARNm finis qui ont subi un coiffage 5', une polyadÊnylation 3' et un Êpissage intron sont exportÊs du noyau vers le cytoplasme. Les prÊ-ARNr et les prÊ-ARN peuvent être traitÊs par clivage intramolÊculaire, Êpissage, mÊthylation et conversion chimique des nuclÊotides. L'Êdition d'ARN est Êgalement effectuÊe, mais rarement, pour insÊrer des bases manquantes après la synthèse d'un ARNm. La traduction
Chaque triplet de nuclÊotides de l'ARN messager indique l'acide aminÊ à incorporer dans la chaÎne polypeptidique qui se forme. La longueur de la protÊine synthÊtisÊe sera donc proportionnelle à la longueur de l'ARNm mature. L'ARNm entier est traduit par Êtapes, codon après codon. Lorsqu'un codon non-sens est rencontrÊ, un facteur de libÊration se lie et dissocie les composants et libère la nouvelle protÊine. Le pliage de la protÊine se produit pendant et après la traduction. Les trois Êtapes principales de la traduction sont : l'initiation, l'Êlongation et la terminaison. L'initiation
ou dĂŠmarrage.
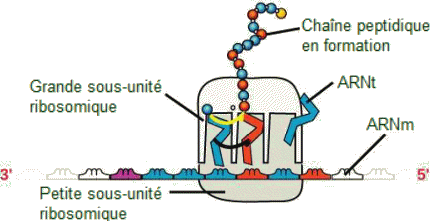
Une fois attachÊe à l'ARNm, cette sous-unitÊ se dÊplace vers l'extrÊmitÊ 3' du messager jusqu'à trouver le codon d'initiation. Un ARNt arrive avec l'anticodon UAC (Met), puis la sous-unitÊ ribosomique principale arrive et, à partir de ce moment, la formation de liaisons peptidiques pourra se produite entre des acides aminÊs sÊquentiels spÊcifiÊs par la matrice d'ARNm selon le code gÊnÊtique. Les ARNt chargÊs pÊnètrent dans le site ribosomal A (Aminoacyl) et leurs acides aminÊs se lient avec l'acide aminÊ au site P (Peptidyl). L'Êlongation.
Ă chaque ĂŠtape,
le mĂŞme scĂŠnario se reproduit : le ribosome lit un nouveau codon de l'ARNm
exposĂŠ sur le site libre A. Un ARNt, disposant Ă une extrĂŠmitĂŠ de l'anticodon
complĂŠmentaire vient s'apparier au codon. L'acide aminĂŠ que cet ARNt
porte Ă son autre extrĂŠmitĂŠ se lie Ă l'acide aminĂŠ apportĂŠ par l'ARNt
prĂŠcĂŠdent et qui est lui-mĂŞme attachĂŠ Ă l'acide aminĂŠ ou aux acides
aminĂŠs dĂŠjĂ liĂŠs entre eux lors des cycles prĂŠcĂŠdents. Cette jonction
(une liaison peptidique) s'effectue grâce à une enzyme prÊsente sur
la grande sous-unitĂŠ du ribosome. L'ARNt peut maintenant se dĂŠtacher
pour laisser libre le site et permettre la lecture du codon suivant et
son appariement avec l'anticodon apportĂŠe par un autre ARNt, etc. Ainsi,
de proche en proche, la sĂŠquence d'acides aminĂŠs grandit-elle jusqu'Ă
l'apparition d'un signal d'arrĂŞt.
La
terminaison.
Les protÊines primaires se replient pour atteindre des structures secondaires, tertiaires ou quaternaires et acquièrent une fonctionnalitÊ biologique. L'expression des
gènes.
Alors que chaque cellule partage le même gÊnome et la même sÊquence d'ADN, chaque cellule n'exprime pas le même ensemble de gènes. Chaque type de cellule a besoin d'un ensemble diffÊrent de protÊines pour remplir sa fonction. Donc, seul un petit sous-ensemble de protÊines est exprimÊ dans une cellule. Pour que les protÊines s'expriment, l'ADN doit être transcrit en ARN et l'ARN doit être traduit en protÊine. Dans un type de cellule donnÊ, tous les gènes codÊs dans l'ADN ne sont pas transcrits en ARN ou traduits en protÊines car des cellules spÊcifiques de notre corps ont des fonctions spÊcifiques. Les protÊines spÊcialisÊes qui composent l'oeil (iris, cristallin et cornÊe) ne sont exprimÊes que dans l'oeil, tandis que les protÊines spÊcialisÊes du coeur (cellules du stimulateur cardiaque, muscle cardiaque et valves) ne sont exprimÊes que dans le coeur. à un moment donnÊ, seul un sous-ensemble de tous les gènes codÊs par notre ADN sont exprimÊs et traduits en protÊines. L'expression de gènes spÊcifiques est un processus hautement rÊgulÊ avec de nombreux niveaux et Êtapes de contrôle. Cette complexitÊ garantit une bonne expression du gène dans la cellule appropriÊe au bon moment. Pour qu'une cellule fonctionne correctement, les protÊines nÊcessaires doivent être synthÊtisÊes au bon moment et au bon endroit. Toutes les cellules contrôlent ou rÊgulent la synthèse des protÊines à partir des informations codÊes dans leur ADN. Le processus d'activation d'un gène pour produire de l'ARN et des protÊines est appelÊ expression gÊnique. Que ce soit dans un organisme unicellulaire simple ou dans un organisme multicellulaire complexe, chaque cellule contrôle quand et comment ses gènes sont exprimÊs. Pour que cela se produise, il doit y avoir des mÊcanismes chimiques internes qui contrôlent quand un gène est exprimÊ pour produire de l'ARN et des protÊines, quelle quantitÊ de protÊines est produite et quand il est temps d'arrêter de produire cette protÊine parce qu'elle n'est plus nÊcessaire. La rÊgulation de l'expression des gènes demanderait une quantitÊ importante d'Ênergie si un un organisme devait exprimer chaque gène à tout moment. Il est donc plus Êconome en Ênergie de n'activer les gènes que lorsqu'ils sont nÊcessaires. De plus, l'expression d'un sous-ensemble de gènes dans chaque cellule permet d'Êconomiser de l'espace car l'ADN doit être dÊroulÊ de sa structure Êtroitement enroulÊe pour transcrire et traduire l'ADN. Les cellules devraient être Ênormes si chaque protÊine Êtait constamment exprimÊe dans chaque cellule. Le contrôle de l'expression des gènes est extrêmement complexe. Les dysfonctionnements de ce processus sont prÊjudiciables à la cellule et peuvent conduire au dÊveloppement de nombreuses maladies, y compris à celui du cancer. L'ARN interfÊrent
et l'intĂŠrfĂŠrence par ARN.
L'ARN
double brin (dsARN).
⢠ARN viral. - De nombreux virus utilisent l'ARN double brin comme matÊriel gÊnÊtique ou le produisent pendant leur cycle de rÊplication. L'interfÊrence par ARN est donc une dÊfense naturelle contre les virus.Les Êtapes clÊs de l'interfÊrence par ARN. Une enzyme appelÊe Dicer reconnaÎt et coupe l'ARN double brin en petits fragments d'environ 20-25 nuclÊotides. Ces petits fragments double brin sont appelÊs petits ARN interfÊrents (siARN) ou microARN (miARN) (selon leur origine et leur mode d'action prÊcis, bien que les miARN soient souvent impliquÊs dans une rÊgulation plus fine et moins dans la dÊgradation directe de l'ARNm comme les siARN). Les siARN sont ensuite chargÊs dans un complexe protÊique appelÊ RISC (RNA-induced silencing complex). L'une des deux brins du siARN (le "brin guide") est conservÊe et incorporÊe dans RISC, tandis que l'autre brin ("brin passager") est gÊnÊralement dÊgradÊe. Une protÊine clÊ de RISC est Argonaute (Ago), qui possède une activitÊ "slicer" (couteau molÊculaire) dans certains cas. Le brin guide du siARN au sein de RISC guide le complexe vers les molÊcules d'ARN messager (ARNm) qui ont une sÊquence complÊmentaire à ce brin guide. L'ARNm est la molÊcule qui porte l'information gÊnÊtique du gène à partir du noyau vers les ribosomes, oÚ les protÊines sont synthÊtisÊes. Une fois que RISC a trouvÊ un ARNm cible, il peut induire le silencieux gÊnique de deux manières principales : ⢠DÊgradation de l'ARNm. - Si la complÊmentaritÊ entre le siARN et l'ARNm est parfaite ou presque parfaite, la protÊine Ago de RISC peut couper (cliver) l'ARNm cible. L'ARNm coupÊ est alors rapidement dÊgradÊ par les enzymes cellulaires. RÊsultat : l'ARNm n'est plus disponible pour la traduction en protÊine, et l'expression du gène correspondant est rÊduite ou complètement supprimÊe. |
| . |
|
|
|||||||||||||||||||||||||||||||
|
