| . |
|
||||||
|
|
| . |
|
||||||
| Les Mollusques |
Les mollusques
sont un embranchement des invertÃĐbrÃĐs.
Ce sont des animaux à corps mou, dÃĐpourvus
de squelette interne; non articulÃĐs, ils sont
ordinairement munis d'une coquille externe ou
interne. Cette coquille peut contenir l'animal en entier; dans certains
cas, elle n'est que rudimentaire, dans quelques autres, elle manque complÃĻtement.
Le systÃĻme nerveux est constituÃĐ par
plusieurs groupes de ganglions reliÃĐs entre
eux par des connectifs; la cavitÃĐ digestive distincte est pourvue de deux
ouvertures; la reproduction est ovipare ou ovovipare. Les Mollusques rassembleraient
quelque 130 000 espÃĻces. C'est l'embranchement le plus riche aprÃĻs celui
des arthropodes. On distingue deux sous-embranchements (ConchifÃĻres et
Amphineures); les trois classes les plus importantes (GastÃĐropodes,
Lamellibranches,
CÃĐphalopodes)
appartiennent au premier.
Avant les Mollusques, les MolluscoÃŊdes. - Dans la classification proposÃĐe par H. Milne-Edwards |
||
| La
systÃĐmatique des mollusques
On rÃĐpartit les mollusques en deux sous-embranchements : les ConchifÃĻres et les Amphineures. Le premier se subdivise en trois grandes classes, ChÃĐphalopodes, GastÃĐropodes et Lamellibranches, auxquelles on ajoute les SolÃĐnoconques et le Monoplacophores. Le second sous-embranchement est formÃĐ de la classe des Chitons (Polyplacophores) et de celle des Aplacophores. |
| ConchifÃĻres | CÃĐphalopodes
(2 sous-classes) :
Dibranchiaux : DÃĐcapodes (seiches, calmars), Octopodes (poulpe ou pieuvre), Vampyromorphes : vampyrotheutis. TÃĐtrabranchiaux : nautiles, ammonites. |
| GastÃĐropodes
Prosobranches : ormeaux, bigorneaux, murex Opisthobranches : liÃĻvres de mer. PulmonÃĐs : limnÃĐes, planorbes, limaces, escargots. |
|
| Lamellibranches
Eulamellibranches : coques, bÃĐnitiers, palourdes. Filibranches : moules, huÃŪtres. Protobranches. Septibranches. |
|
| SolÃĐnoconques (ou Scaphopodes) : dentale. | |
| Monoplacophores (rares aujourd'hui, surtout fossiles). | |
| Amphineures | Polyplacophores
(ou Chitons)
Acanthochitons : chiton polii. Ischinichitons LÃĐpidopleures : lepidopleurus cancellatus. |
| Aplacophores
CaudofovÃĐates : chaetodermes, limifossor SolÃĐnogastres : pruvotina, epimenia, nematodermia. |
| Les
ConchifÃĻres.
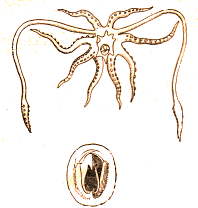
Les CÃĐphalopodes. La classe des CÃĐphalopodes reprÃĐsente le sommet de l'ÃĐvolution des mollusques en termes de complexitÃĐ anatomique, de dÃĐveloppement sensoriel et de comportement. Exclusivement marins, ces prÃĐdateurs sophistiquÃĐs ont conquis tous les ocÃĐans, des rÃĐcifs coralliens tropicaux aux abysses les plus profonds. Leur histoire ÃĐvolutive est ancienne et prestigieuse, remontant au Cambrien supÃĐrieur, il y a plus de 500 millions d'annÃĐes. Leur ÃĒge d'or se situe au PalÃĐozoÃŊque et au MÃĐsozoÃŊque, oÃđ ils rÃĐgnaient en maÃŪtres sur les mers sous la forme d'une incroyable diversitÃĐ d'ammonites, de bÃĐlemnites et de nautiloÃŊdes. La grande majoritÃĐ de ces formes à coquille externe a disparu lors de l'extinction CrÃĐtacÃĐ-PalÃĐogÃĻne, laissant la place aux formes modernes à coquille interne rÃĐduite ou absente, comme les pieuvres, les calmars et les seiches. Aujourd'hui, on compte environ 800 espÃĻces actuelles, classÃĐes principalement en deux sous-classes : les NautiloÃŊdes, vÃĐritables fossiles vivants avec leur coquille externe cloisonnÃĐe, et les ColÃĐoÃŊdes, qui regroupent la quasi-totalitÃĐ des espÃĻces familiÃĻres. Le plan d'organisation
fondamental d'un cÃĐphalopode est une variation extrÊme du modÃĻle ancestral
des mollusques, orientÃĐ vers une vie pÃĐlagique active et la prÃĐdation.
Le corps se compose d'une tÊte proÃĐminente, portant des yeux remarquablement
complexes et une couronne de bras musculeux et prÃĐhensiles, et d'un sac
viscÃĐral allongÃĐ contenant les organes. Le pied du mollusque primitif
s'est diffÃĐrenciÃĐ en deux structures : l'entonnoir (ou siphon), un tube
musculeux ventral utilisÃĐ pour la propulsion par rÃĐaction, et la couronne
de bras et de tentacules qui entourent la bouche. Ces appendices, munis
de ventouses parfois armÃĐes de crochets cornÃĐs, servent à la capture
des proies, à la manipulation d'objets et à la locomotion sur le fond.
Chez les calmars et les seiches, huit bras courts et deux longs tentacules
rÃĐtractiles, terminÃĐs par une massue, permettent une attaque ÃĐclair.
Chez les pieuvres, seuls huit bras de longueur ÃĐgale existent, explorant
chaque anfractuositÃĐ du substrat.
Loligo vulgaris L. (bras et bouche). La locomotion est une spÃĐcialitÃĐ des cÃĐphalopodes. La propulsion par rÃĐaction est leur mode de dÃĐplacement le plus spectaculaire : l'eau est aspirÃĐe dans la cavitÃĐ pallÃĐale puis violemment expulsÃĐe par l'entonnoir orientable, permettant des accÃĐlÃĐrations fulgurantes, que ce soit pour chasser ou pour fuir. Ce systÃĻme est couplÃĐ Ã des nageoires latÃĐrales ondulantes chez les calmars et les seiches, offrant une nage stationnaire, une maniabilitÃĐ et une ÃĐconomie d'ÃĐnergie hors pair. Les pieuvres, quant à elles, sont principalement benthiques et se dÃĐplacent en rampant sur leurs bras ou, plus rarement, par propulsion. La flottabilitÃĐ est un autre dÃĐfi magistralement rÃĐsolu, notamment par la seiche avec son os de seiche : une coquille interne poreuse et cloisonnÃĐe dont les chambres sont remplies d'un mÃĐlange de gaz et de liquide dont les proportions sont finement rÃĐgulÃĐes par un tissu spÃĐcialisÃĐ, le siphoncule, pour maintenir une flottaison neutre à n'importe quelle profondeur. Le nautile utilise le mÊme principe dans sa coquille externe, tandis que certains calmars des profondeurs accumulent des ions ammonium, plus lÃĐgers que l'eau de mer, dans leurs tissus. Le systÃĻme nerveux et les organes des sens des cÃĐphalopodes n'ont pas d'ÃĐquivalent chez les autres invertÃĐbrÃĐs et rivalisent avec ceux des vertÃĐbrÃĐs. Le cerveau, protÃĐgÃĐ par une capsule cartilagineuse, est le rÃĐsultat de la fusion de plusieurs ganglions nerveux et prÃĐsente des lobes spÃĐcialisÃĐs dans l'apprentissage et la mÃĐmoire. Les yeux, en particulier, sont un exemple frappant d'ÃĐvolution convergente avec ceux des poissons : ils possÃĻdent une cornÃĐe, un cristallin, un iris et une rÃĐtine. Contrairement à l'Åil des vertÃĐbrÃĐs, les fibres nerveuses partent de l'arriÃĻre de la rÃĐtine, ÃĐvitant le point aveugle. Les colÃĐoÃŊdes peuvent se focaliser en dÃĐplaçant le cristallin d'avant en arriÃĻre, comme un objectif d'appareil photo, et perçoivent mÊme la polarisation de la lumiÃĻre. Ils sont nÃĐanmoins gÃĐnÃĐralement daltoniens. La sensorialitÃĐ ne s'arrÊte pas là : leur peau tout entiÃĻre est un organe sensoriel photosensible grÃĒce à des protÃĐines, les opsines, leur donnant une perception de la lumiÃĻre et peut-Être des motifs environnants indÃĐpendamment des yeux. Des organes analogues à l'oreille interne, les statocystes, assurent un sens de l'ÃĐquilibre et de l'orientation d'une prÃĐcision remarquable. Leur comportement est à la hauteur de leur ÃĐquipement neuronal. Ce sont des prÃĐdateurs insatiables, traquant et capturant crustacÃĐs, poissons et autres mollusques avec des stratÃĐgies de chasse ÃĐlaborÃĐes, incluant la discrÃĐtion, l'affÃŧt ou la poursuite active. Un bec cornÃĐe et puissant, semblable à celui d'un perroquet mais inversÃĐ, leur permet de dÃĐchiqueter leurs proies, aidÃĐ par une rÃĒpe linguale, la radula. Une glande digestive volumineuse complÃĻte un systÃĻme digestif performant. La communication et le camouflage sont poussÃĐs à un degrÃĐ inouÃŊ grÃĒce à un systÃĻme tÃĐgumentaire unique. Des millions de cellules spÃĐcialisÃĐes dans la peau, les chromatophores (contenant des pigments), les iridophores (rÃĐflÃĐchissant la lumiÃĻre) et les leucophores (diffusant la lumiÃĻre ambiante), sont sous contrÃīle nerveux direct. Cela permet à un animal comme la seiche de modifier la couleur, le motif, la texture et la rÃĐflectance de sa peau en une fraction de seconde, crÃĐant des affichages hypnotiques pour communiquer avec ses congÃĐnÃĻres, intimider un rival ou se fondre dans le dÃĐcor avec un mimÃĐtisme quasi parfait. Le nuage d'encre, expulsÃĐ par l'entonnoir, est un leurre ultime, crÃĐant soit un ÃĐcran opaque (pseudomorphe), soit une nuÃĐe de petites gouttelettes qui imitent leur propre silhouette pour tromper l'agresseur. La biologie de la reproduction est tout aussi singuliÃĻre. Les sexes sont sÃĐparÃĐs et le dimorphisme sexuel peut Être marquÃĐ. Un des bras du mÃĒle, l'hectocotyle, est modifiÃĐ pour collecter les spermatophores (paquets de spermatozoÃŊdes) et les transfÃĐrer dans la cavitÃĐ pallÃĐale de la femelle, parfois en se dÃĐtachant du corps du mÃĒle lors d'un accouplement à distance, un phÃĐnomÃĻne qui lui a valu d'Être d'abord dÃĐcrit comme un ver parasite. AprÃĻs l'accouplement et la ponte d'Åufs gÃĐnÃĐralement gros et riches en vitellus, souvent fixÃĐs en grappes au substrat, la femelle veille et nettoie sa ponte sans relÃĒche jusqu'à l'ÃĐclosion. La reproduction est sÃĐmelpare : cette garde acharnÃĐe est la derniÃĻre fonction de l'animal, qui cesse de s'alimenter, subit un vieillissement programmÃĐ catastrophique et meurt peu aprÃĻs l'ÃĐclosion des jeunes. Ces derniers, minuscules rÃĐpliques des adultes, adoptent immÃĐdiatement un mode de vie planctonique ou benthique, sans aucun soin parental ultÃĐrieur. Ce cycle de vie, combinÃĐ Ã une croissance exceptionnellement rapide, fait des cÃĐphalopodes des acteurs ÃĐcologiques majeurs, dont les prolifÃĐrations sporadiques peuvent avoir des impacts considÃĐrables sur les chaÃŪnes alimentaires marines. Les
GastÃĐropodes.
Le plan d'organisation qui dÃĐcoule de cette torsion, puis des ÃĐvolutions subsÃĐquentes, s'articule autour de trois grandes rÃĐgions. La tÊte est bien individualisÃĐe et porte typiquement une ou deux paires de tentacules sensoriels, des yeux dont la complexitÃĐ varie d'une simple fossette pigmentaire à une vÃĐritable vÃĐsicule optique avec cristallin, et une bouche ventrale. La cavitÃĐ buccale abrite un organe caractÃĐristique, la radula, une rÃĒpe chitineuse constituÃĐe de milliers de dents microscopiques, dont la forme extraordinairement variable (en aiguillon, en lame, en harpon, voire perdue) reflÃĻte le rÃĐgime alimentaire, qu'il soit herbivore, carnivore, charognard, suspensivore ou parasite. Le pied, large sole rampante ventrale, est l'organe locomoteur par excellence. Sa surface est parcourue d'ondes de contractions musculaires directes ou rÃĐtrogrades, lubrifiÃĐes par un mucus sÃĐcrÃĐtÃĐ par une glande pÃĐdieuse, permettant une locomotion par reptation d'une efficacitÃĐ remarquable, mÊme sur des surfaces verticales ou lisses. Chez les formes pÃĐlagiques, le pied peut se transformer en une nageoire ondulante ou en deux ailerons, les ÂŦ pieds de papillon Âŧ des ptÃĐropodes. La masse viscÃĐrale est, chez la majoritÃĐ des espÃĻces, enroulÃĐe en spirale et contenue dans une coquille univalve. Cette coquille, sÃĐcrÃĐtÃĐe par le manteau et typiquement conique, s'enroule selon un axe, la columelle. L'enroulement peut Être planispiralÃĐ (trÃĻs rare) ou, le plus souvent, hÃĐlicoÃŊdal (en forme de vis), rÃĐsultant en un cÃīne compact et solide. L'immense diversitÃĐ des formes de coquilles, des spires allongÃĐes des aiguillettes aux formes globuleuses des buccins, en passant par les porcelaines polies, constitue un champ d'ÃĐtude en soi. Pourtant, une tendance ÃĐvolutive majeure et rÃĐcurrente chez les gastÃĐropodes est la rÃĐduction, l'internalisation ou la perte totale de la coquille, phÃĐnomÃĻne observÃĐ indÃĐpendamment dans de trÃĻs nombreuses lignÃĐes, donnant naissance aux diverses "limaces", marines ou terrestres. L'adaptation aux diffÃĐrents milieux a profondÃĐment modifiÃĐ l'appareil respiratoire. Les gastÃĐropodes aquatiques, comme les patelles ou les bigorneaux, respirent grÃĒce à des branchies (ctÃĐnidies) situÃĐes dans la cavitÃĐ pallÃĐale, gÃĐnÃĐralement une seule du cÃītÃĐ gauche suite à la torsion et aux asymÃĐtries compensatoires. Chez les gastÃĐropodes terrestres (escargots et limaces) et certains dulçaquicoles comme les planorbes, la cavitÃĐ pallÃĐale s'est transformÃĐe en un poumon richement vascularisÃĐ. Ces pulmonÃĐs ont perdu leurs branchies et respirent l'air atmosphÃĐrique par un orifice contractile, le pneumostome, qui s'ouvre et se ferme pÃĐriodiquement. Cette conquÊte du milieu aÃĐrien s'est accompagnÃĐe d'autres adaptations cruciales, comme l'excrrÃĐtion d'acide urique pour ÃĐconomiser l'eau et une reproduction souvent hermaphrodite. Le systÃĻme nerveux des gastÃĐropodes, bien que moins centralisÃĐ que celui des cÃĐphalopodes, est typiquement gangliÃĐnaire, avec un anneau pÃĐri-Åsophagien et des connectifs vers les ganglions pÃĐdieux et viscÃĐraux. Chez les formes à torsion, les connectifs nerveux forment une figure en huit caractÃĐristique. Certains neurones gÃĐants identifiables d'une espÃĻce à l'autre, notamment chez l'aplysie ou la limnÃĐe, ont fait de ces animaux des modÃĻles inestimables en neurobiologie pour l'ÃĐtude des bases cellulaires de l'apprentissage et de la mÃĐmoire. La biologie sensorielle et comportementale des gastÃĐropodes est bien plus riche que leur apparente lenteur ne le suggÃĻre. Les tentacules et le bord du manteau sont des zones de grande sensibilitÃĐ tactile et chimique. L'osphradie, un organe sensoriel spÃĐcialisÃĐ situÃĐ dans la cavitÃĐ pallÃĐale, fonctionne comme un chimiorÃĐcepteur capable d'analyser la qualitÃĐ de l'eau. La vision est gÃĐnÃĐralement sommaire mais peut Être ÃĐtonnamment dÃĐveloppÃĐe chez certains prÃĐdateurs comme les cÃīnes, dont les yeux sont de vÃĐritables chambres noires. Les comportements de prÃĐdation, de fuite ou de reproduction peuvent Être d'une complexitÃĐ inattendue. Les cÃīnes sont des chasseurs redoutables, capables d'injecter un cocktail de conotoxines paralysantes à l'aide d'une dent radulaire modifiÃĐe en harpon empoisonnÃĐ, un venin assez puissant pour tuer un humain. La reproduction offre un tableau presque infini de variations. Si la plupart des gastÃĐropodes marins sont gonochoriques (sexes sÃĐparÃĐs) avec une fÃĐcondation externe conduisant à une larve vÃĐligÃĻre planctonique, de nombreuses espÃĻces ont une fÃĐcondation interne et pondent des Åufs protÃĐgÃĐs dans des capsules cornÃĐes fixÃĐes au substrat. Les pulmonÃĐs sont presque exclusivement hermaphrodites, avec un appareil reproducteur d'une complexitÃĐ anatomique extraordinaire, incluant souvent un organe copulateur, une bourse copulatrice et des glandes annexes produisant les diffÃĐrentes enveloppes de l'oeuf. L'accouplement rÃĐciproque est la rÃĻgle, souvent prÃĐcÃĐdÃĐ de parades nuptiales spectaculaires. Le cÃĐlÃĻbre "coup de dard d'amour" de certains escargots terrestres, oÃđ un dard calcaire est enfoncÃĐ dans le corps du partenaire pour transfÃĐrer des hormones stimulant la reproduction, reste l'un des rituels les plus singuliers du rÃĻgne animal. Cette immense diversitÃĐ anatomique, ÃĐcologique et comportementale fait des gastÃĐropodes un groupe d'ÃĐtude central en biologie ÃĐvolutive, en ÃĐcologie des communautÃĐs et en biologie du dÃĐveloppement, tout en ÃĐtant un maillon essentiel de presque tous les ÃĐcosystÃĻmes, du recyclage de la matiÃĻre organique à la rÃĐgulation des populations de leurs proies. Les
Lamellibranches.
Le mode de vie sÃĐdentaire ou fixÃĐ a entraÃŪnÃĐ une profonde redÃĐfinition des fonctions vitales. La locomotion est gÃĐnÃĐralement rÃĐduite. Chez les formes fouisseuses, comme les couteaux ou les palourdes, le pied musculeux, comprimÃĐ en forme de hache, s'engorge de sang et s'ancre dans le sÃĐdiment avant que sa contraction ne tire l'animal vers le bas. Chez les espÃĻces fixÃĐes comme les moules, le pied est atrophiÃĐ et sert principalement à sÃĐcrÃĐter un byssus, un faisceau de filaments cornÃĐs qui ancre solidement l'animal au substrat. Les huÃŪtres, elles, sont cimentÃĐes au rocher par une sÃĐcrÃĐtion directe de la valve gauche. La locomotion peut Être plus dynamique chez certaines espÃĻces libres comme les pÃĐtoncles, qui pratiquent une nage par rÃĐaction en expulsant violemment l'eau de leur cavitÃĐ en claquant rapidement leurs valves. L'alimentation et la respiration sont intimement liÃĐes et reposent sur un systÃĻme de filtration d'une efficacitÃĐ prodigieuse. La cavitÃĐ pallÃĐale est traversÃĐe par un courant d'eau continu, crÃĐÃĐ par le battement de cils vibratiles. L'eau entre par un siphon inhalant, souvent prolongÃĐ par un tube charnu, passe au travers de branchies extraordinairement modifiÃĐes, et ressort par un siphon exhalant. Les branchies, ou ctÃĐnidies, ne sont plus de simples organes respiratoires mais sont devenues de vastes lames filtrantes plissÃĐes (d'oÃđ le nom de la classe), recouvertes d'un tapis de cils et d'un mucus collant. Les particules alimentaires en suspension (phytoplancton, bactÃĐries, dÃĐtritus organiques) sont piÃĐgÃĐes par ce mucus, acheminÃĐes par des gouttiÃĻres ciliÃĐes vers les palpes labiaux, triÃĐes, puis portÃĐes à la bouche. Ce systÃĻme permet à un banc d'huÃŪtres de filtrer l'ÃĐquivalent d'un grand volume d'eau en une seule journÃĐe, jouant un rÃīle ÃĐcologique fondamental dans la clarification de l'eau et le transfert d'ÃĐnergie du plancton vers le benthos. Le tube digestif est simplifiÃĐ, sans radula, et un stylet cristallin, une tige gÃĐlatineuse hyaline, tourne dans l'estomac pour libÃĐrer des enzymes digestives et brasser les aliments. La reproduction est gÃĐnÃĐralement gonochorique, avec une fÃĐcondation externe et le passage par une larve vÃĐligÃĻre planctonique, qui possÃĻde dÃĐjà une ÃĐbauche de coquille. Un cas fascinant est celui des moules d'eau douce de l'ordre des Unionoida, dont la larve, le glochidium, est un parasite temporaire des branchies ou de la peau des poissons, assurant ainsi sa dispersion vers l'amont des cours d'eau, une stratÃĐgie vitale pour un animal adulte sÃĐdentaire. Les
SolÃĐnoconques.
Exclusivement marins et benthiques, ils vivent enfouis verticalement dans les sÃĐdiments meubles, depuis la zone littorale jusqu'à des profondeurs abyssales, laissant dÃĐpasser à la surface du sable uniquement l'extrÃĐmitÃĐ la plus fine de leur coquille, le pavillon apical. L'animal est entiÃĻrement contenu dans ce tube, et son plan d'organisation est une rÃĐponse adaptative parfaite à la vie interstitielle. La tÊte est rÃĐduite, sans yeux, mais prolongÃĐe par un proboscis. La radula est prÃĐsente, et surtout, elle est entourÃĐe de deux touffes de fins tentacules prÃĐhensiles et ciliÃĐs, les captacules. Ces derniers, sortant par la grande ouverture antÃĐrieure de la coquille, explorent les espaces interstitiels du sÃĐdiment environnant. Lorsqu'un captacule rencontre une proie microscopique, un foraminifÃĻre par exemple, il y adhÃĻre et la ramÃĻne vers la bouche. Les solÃĐnoconques sont donc des microcarnivores sÃĐlectifs, se nourrissant de la microfaune du sable. Le pied, conique et musculeux, sort de l'ouverture antÃĐrieure et sert à l'enfouissement. Sa forme effilÃĐe et sa capacitÃĐ Ã se dilater à son extrÃĐmitÃĐ par pression sanguine lui permettent de creuser efficacement dans le sÃĐdiment en agissant comme une ancre et une pelle. La respiration, en l'absence de branchies typiques, est assurÃĐe par la surface mÊme de la cavitÃĐ pallÃĐale, un manteau en forme de sac interne parcouru de cils. L'eau n'entre pas par des siphons mais est mise en mouvement par les battements ciliaires du manteau, entrant et sortant par la mÊme petite ouverture apicale, au sommet de la coquille. Ce flux unidirectionnel, aspirant par l'apex et rejetÃĐ par l'apex, sert à la fois à la respiration et à l'ÃĐvacuation des dÃĐchets, puisque l'anus dÃĐbouche dans cette cavitÃĐ pallÃĐale. La sexualitÃĐ est distincte, et la fÃĐcondation externe donne naissance à une larve trochophore, puis vÃĐligÃĻre, caractÃĐristique des mollusques, qui mÃĻne une vie planctonique avant de se mÃĐtamorphoser et de gagner le fond. Longtemps classÃĐs par dÃĐfaut entre les gastÃĐropodes et les lamellibranches, les scaphopodes forment une lignÃĐe clairement individualisÃĐe par leur combinaison unique de caractÃĻres : une coquille tubulaire, une alimentation par captacules, une respiration pallÃĐale non branchiale et une adaptation trÃĻs poussÃĐe à la microfaune interstitielle, faisant d'eux un groupe discret mais au succÃĻs ÃĐvolutif constant. Les
Monoplacophores.
Sur le plan morphologique, les monoplacophores prÃĐsentent une anatomie singuliÃĻre qui tranche avec celle de la plupart des autres mollusques. Comme leur nom l'indique, ils sont protÃĐgÃĐs par une coquille unique, en forme de capuchon ou de petite patelle, qui recouvre l'ensemble de leur masse corporelle. Sous cette coquille, le manteau dÃĐlimite une large cavitÃĐ pallÃĐale. L'une de leurs caractÃĐristiques les plus remarquables est leur symÃĐtrie bilatÃĐrale stricte, contrastant avec l'asymÃĐtrie souvent observÃĐe chez les gastÃĐropodes due à la torsion. De plus, ils prÃĐsentent une organisation interne trÃĻs particuliÃĻre caractÃĐrisÃĐe par une rÃĐpÃĐtition sÃĐrielle de certains organes. On observe ainsi plusieurs paires de branchies (ctÃĐnidies), de nÃĐphridies (organes excrÃĐteurs), de gonades et de muscles rÃĐtracteurs de la coquille disposÃĐs de part et d'autre de l'axe du corps. Cette pseudo-mÃĐtamÃĐrie a longtemps fait croire qu'ils reprÃĐsentaient un chaÃŪnon manquant entre les mollusques et les annÃĐlides, bien que les ÃĐtudes phylogÃĐnÃĐtiques modernes les classent fermement au sein de l'embranchement des mollusques, probablement proches des aplacophores ou à la base des conchifÃĻres. Les monoplacophores sont des animaux des grands fonds, ÃĐvoluant gÃĐnÃĐralement dans les plaines abyssales, entre 1800 et plus de 6000 mÃĻtres de profondeur, sur des fonds meubles. Leur large pied musculeux, situÃĐ sur la face ventrale, leur permet de ramper lentement sur la vase. Ils se nourrissent principalement en broutant les micro-organismes et les dÃĐbris organiques prÃĐsents dans les sÃĐdiments, à l'aide de leur radula, une langue rÃĒpeuse typique des mollusques. En raison de leur habitat extrÊme, difficile d'accÃĻs, les monoplacophores restent rares et mal connus, mais leur ÃĐtude continue d'apporter des ÃĐclairages prÃĐcieux sur la morphologie ancestrale des mollusques et l'adaptation de la vie aux environnements abyssaux. Les Amphineures.
Leur anatomie est entiÃĻrement dictÃĐe par une vie benthique sur les substrats durs, principalement dans la zone intertidale, oÃđ leur adaptation à l'ÃĐcrasement par les vagues est poussÃĐe à l'extrÊme. La coquille à huit valves, enchÃĒssÃĐe dans une ceinture musculaire charnue, la ceinture, permet à l'animal d'ÃĐpouser les irrÃĐgularitÃĐs de la roche tout en offrant une protection solide, et surtout la capacitÃĐ unique de s'enrouler en boule, à la maniÃĻre d'un cloporte, lorsqu'il est dÃĐlogÃĐ. Le corps, de forme ovale et aplatie dorso-ventralement, ne montre pas de tÊte distincte : les yeux et les tentacules cÃĐphaliques sont absents, la face dorsale ÃĐtant entiÃĻrement recouverte par l'armure et la ceinture, qui peut Être lisse ou ornÃĐe d'ÃĐpines, de soies ou d'ÃĐcailles calcaires. Sous les valves, la face ventrale est entiÃĻrement dominÃĐe par un pied large, plat et musculeux, organe locomoteur et d'adhÃĐrence formidable. Ce pied, couplÃĐ Ã la succion produite par le manteau qui adhÃĻre ÃĐtroitement au substrat, permet au chiton de rÃĐsister aux forces de cisaillement des vagues les plus puissantes. De part et d'autre du pied s'ÃĐtend la cavitÃĐ pallÃĐale, un sillon profond dans lequel sont logÃĐes en rangÃĐes les branchies, les ctÃĐnidies, dont le nombre varie de quelques unitÃĐs à plusieurs dizaines de paires, une caractÃĐristique primitive et un trait diagnostique du groupe. L'alimentation est principalement assurÃĐe par une radula extrÊmement robuste et minÃĐralisÃĐe, l'une des plus dures du rÃĻgne animal, dont les dents latÃĐrales sont imprÃĐgnÃĐes de magnÃĐtite, un minÃĐral ferreux. Cette radula magnÃĐtique leur permet de rÃĒper avec une efficacitÃĐ exceptionnelle les algues incrustantes, les diatomÃĐes et mÊme le biofilm bactÃĐrien sur les rochers. Certaines espÃĻces sont devenues des prÃĐdateurs actifs, soulevant le bord de leur manteau pour piÃĐger de petits crustacÃĐs ou vers sous leur pied. Le systÃĻme nerveux est typiquement amphineure et tÃĐtraneure, c'est-à -dire qu'il ne forme pas de ganglions concentrÃĐs mais consiste en un anneau nerveux pÃĐri-oesophagien d'oÃđ partent quatre cordons nerveux longitudinaux (deux pÃĐdieux et deux pallÃĐo-viscÃĐraux) reliÃĐs entre eux par des commissures transversales, comme une ÃĐchelle de corde. Ce caractÃĻre primitif s'accompagne d'organes sensoriels originaux : les esthÃĻtes (ou aesthÃĻtes). Ce sont des canaux sensoriels qui traversent les valves calcaires et dÃĐbouchent à leur surface. Ils contiennent des cellules photorÃĐceptrices et des mÃĐcanorÃĐcepteurs, offrant au chiton une perception diffuse de la lumiÃĻre et du toucher à travers sa propre coquille, un cas unique chez les mollusques. Parfois, ces esthÃĻtes se transforment en de vÃĐritables petits yeux, avec cristallin et rÃĐtine, dispersÃĐs par milliers sur la surface des valves. La reproduction est gonochorique, avec fÃĐcondation externe et ÃĐmission en pleine eau des gamÃĻtes, donnant naissance à une larve trochophore planctonique. Certaines espÃĻces, notamment arctiques, montrent une forme de soin parental rare, la femelle couvant ses Åufs dans sa cavitÃĐ pallÃĐale. Le dÃĐveloppement amÃĻne à une mÃĐtamorphose progressive oÃđ la jeune coquille à sept plaques et le pied se dÃĐveloppent, avant que le jeune chiton ne rejoigne le benthos. Les
Aplacophores.
On distingue traditionnellement deux lignÃĐes : les SolÃĐnogastres, qui vivent gÃĐnÃĐralement en rampant sur les cnidaires, notamment les coraux et les gorgones, dont ils broutent les polypes, et les CaudofovÃĐates, fouisseurs dans les sÃĐdiments vaseux, se nourrissant de foraminifÃĻres et de micro-organismes. La caractÃĐristique fondamentale des Aplacophores est l'absence totale de coquille, mais aussi la rÃĐduction ou la perte du pied rampant caractÃĐristique des mollusques. Chez les CaudofovÃĐates, le pied est presque inexistant; chez les SolÃĐnogastres, il est rÃĐduit à une ÃĐtroite gouttiÃĻre ventrale ciliÃĐe, la gouttiÃĻre pÃĐdieuse, qui se termine parfois par un repli permettant une reptation limitÃĐe. La face ventrale est entiÃĻrement parcourue par ce sillon ciliÃĐ, organe de locomotion discret, trÃĻs ÃĐloignÃĐ du pied large et musculeux des chitons ou des escargots. La tÊte est peu ou pas individualisÃĐe, sans yeux ni tentacules, portant simplement la bouche. La cavitÃĐ buccale contient une radula prÃĐsente chez la plupart des espÃĻces, souvent rÃĐduite à une rangÃĐe de dents, parfois absente, qui sert à racler les proies. Chez les prÃĐdateurs de cnidaires, la radula et les glandes salivaires sont adaptÃĐes pour ingÃĐrer les tissus mous sans dÃĐclencher les nÃĐmatocystes dÃĐfensifs. Une particularitÃĐ frappante est la prÃĐsence d'un organe sensoriel vestibulaire antÃĐrieur, propre aux SolÃĐnogastres, probablement impliquÃĐ dans la chimiorÃĐception et la dÃĐtection des proies. La respiration se fait à travers la surface du corps, puisqu'il n'existe pas de branchies diffÃĐrenciÃĐes, à l'exception des CaudofovÃĐates qui possÃĻdent une paire de ctÃĐnidies bipectinÃĐes dans une petite cavitÃĐ pallÃĐale postÃĐrieure, vestige de l'ancÊtre commun. Le systÃĻme nerveux est amphineure, semblable à une ÃĐchelle de corde, sans ganglions cÃĐrÃĐbroÃŊdes distincts, bien que des ganglions antÃĐrieurs aient ÃĐtÃĐ identifiÃĐs, prÃĐfigurant la cÃĐphalisation des autres mollusques. La reproduction des Aplacophores est sexuÃĐe, les sexes ÃĐtant gÃĐnÃĐralement sÃĐparÃĐs; la fÃĐcondation est externe ou interne, et le dÃĐveloppement mÃĻne à une larve trochophore libre, qui se mÃĐtamorphose en adulte en acquÃĐrant ses spicules. L'ÃĐtude des Aplacophores est importante pour comprendre les origines du plan d'organisation des mollusques. Leur corps vermiforme et l'absence de coquille ont longtemps ÃĐtÃĐ interprÃĐtÃĐs comme une condition ancestrale, suggÃĐrant que les premiers mollusques ressemblaient à un ver recouvert d'une cuticule à spicules, avant l'apparition de la coquille dorsale et du pied musculeux. Aujourd'hui, leur position phylogÃĐnÃĐtique dÃĐbattue les place comme un tÃĐmoignage vivant des formes transitionnelles qui ont jalonnÃĐ la radiation de l'un des phylums les plus inventifs du rÃĻgne animal. |
||
| L'organisme
des mollusques
SystÃĻme nerveux.
1° aux ganglions ventraux, lesquels fournissent les nerfs aux organes de la respiration, aux viscÃĻres et au manteau, constituant ainsi les ganglions branchiaux, sous-oesophagiens, viscÃĐraux;Chez les Lamellibranches le systÃĻme nerveux est trÃĻs simplifiÃĐ. On remarque les ganglions cÃĐrÃĐbroÃŊdes rÃĐunis par une commissure assez dÃĐveloppÃĐe dans certains groupes, abrÃĐgÃĐe dans d'autres, enfin presque rudimentaires chez quelques-uns. Les ganglions pÃĐdieux sont rÃĐunis aux cÃĐrÃĐbroÃŊdes par une paire de connectifs formant un anneau d'autant plus ÃĐtendu que le pied se trouve plus ÃĐcartÃĐ de la bouche. Chez les HuÃŪtres et chez quelques autres genres qui sont dÃĐpourvus de pied les ganglions pÃĐdieux sont atrophiÃĐs. Les ganglions branchiaux sont rÃĐunis aux cÃĐrÃĐbroÃŊdes par des connectifs trÃĻs longs; ces ganglions donnent naissance aux nerfs des muscles adducteurs, postÃĐrieurs des valves, aux filets nerveux des siphons, aux nerfs pallÃĐaux, etc. SystÃĻme circulatoire.
Organes de la
respiration.
Organes reproducteurs.
L'organe reproducteur des GastÃĐropodes hermaphrodites (PulmonÃĐs) est composÃĐ comme il suit : les orifices gÃĐnitaux confondus s'ouvrent dans un seul cloaque : dans ce cas, l'appareil est constituÃĐ par une glande lobulÃĐe dont les follicules portent les ovules à l'extÃĐrieur et les cellules spermatiques à l'intÃĐrieur, et la glande hermaphrodite est gÃĐnÃĐralement nommÃĐe organe en grappe; par un canal excrÃĐteur de l'organe en grappe, tortueux, atteignant l'extrÃĐmitÃĐ postÃĐrieure du canal dÃĐfÃĐrent et de la matrice, tout auprÃĻs de la glande albuminipare, glande en forme de langue, d'un blanc jaunÃĒtre. Elle sÃĐcrÃĻte un liquide ÃĐpais dans lequel nagent des globules et des granulations; son canal excrÃĐteur s'ouvre dans la partie postÃĐrieure de la matrice : cette derniÃĻre a la forme d'un canal un peu tortueux, à parois boursouflÃĐes; elle est adhÃĐrente dans tout son trajet au canal dÃĐfÃĐrent. La matrice en se sÃĐparant du canal dÃĐfÃĐrent forme le vagin, lequel aboutit au vestibule on poche commune. Le vestibule a la forme d'un sac plus ou moins ÃĐtendu; il contient les orifices du vagin, du canal de la poche copulatrice, de la verge; son orifice externe est placÃĐ en arriÃĻre du grand tentacule droit. Le canal dÃĐfÃĐrent est partagÃĐ en deux parties distinctes : la premiÃĻre postÃĐrieure, adhÃĐrente au bord concave de la matrice; l'intÃĐrieure libre. La verge est contenue dans un sac à peu prÃĻs cylindrique; elle passe dans le vestibule, lequel se referme lors de l'accouplement. Le vagin est souvent accompagnÃĐ d'organes accessoires ordinairement rameux, allongÃĐs et rÃĐunis par groupes : ils portent le nom de vÃĐsicules multifides. Dans certaines espÃĻces (Helix pomatia, vermiculata), les branches de chaque vÃĐsicule sont nombreuses; dans quelques autres, il n'existe qu'une seule vÃĐsicule. Ces vÃĐsicules multifides n'existent pas chez les espÃĻces des genres Daudebardia et Zonites; elles sont reprÃĐsentÃĐes par un revÊtement glandulaire considÃĐrÃĐ comme une prostate vaginale. Tous les CÃĐphalopodes sont dioÃŊques : les mÃĒles sont plus petits que les femelles. L'appareil de la reproduction femelle est constituÃĐ par un ovaire enveloppÃĐ par une tunique particuliÃĻre; l'oviducte, continuation de cette tunique, est parfois simple et dÃĐbouche dans le cloaque vers la base de l'entonnoir, parfois double, et, dans ce cas, leurs orifices sont situÃĐs entre les branchies et le rectum. La majeure partie des Mollusques sont ovipares; quelques-uns sont ovovivipares; ces derniers sont en trÃĻs petit nombre. Chez ces animaux la fÃĐconditÃĐ est particuliÃĻrement, dÃĐveloppÃĐe. Poli dÃĐclare que l'HuÃŪtre comestible peut pondre en une seule saison 120 000 oeufs; d'aprÃĻs Davaine, les oeufs pondus par une seule HuÃŪtre atteindraient le chiffre de 4 250 000 oeufs, l'Unio pictorum donnerait 220 000 oeufs. Les PulmonÃĐs sont loin d'atteindre un chiffre aussi ÃĐlevÃĐ. Ces derniers dÃĐposent leurs oeufs dans les lieux frais, sous les mousses, sous les pierres, souvent dans la terre, en un trou creusÃĐ par l'animal et dont la profondeur peut atteindre de 4 à 40 cm. L'ÃĐclosion, d'aprÃĻs Moquin-Tandon commence vers le quinziÃĻme ou le vingtiÃĻme jour, et, dans certains cas, seulement un mois aprÃĻs la ponte. (J. Mabille). |
| . |
|
|
|||||||||||||||||||||||||||||||
|
