 |
La mitose est
un processus de division
cellulaire permettant à une cellule mÃĻre de donner naissance
à deux cellules filles gÃĐnÃĐtiquement identiques. Ce mÃĐcanisme assure
la croissance, la rÃĐgÃĐnÃĐration
et le renouvellement des tissus chez les organismes
multicellulaires. Il se dÃĐroule en plusieurs ÃĐtapes successives impliquant
des modifications structurales du noyau et du cytoplasme.
D'abord, la cellule rÃĐplique son matÃĐriel
gÃĐnÃĐtique afin de disposer de deux copies complÃĻtes de son ADN.
Ensuite, la chromatine se condense pour former
des chromosomes visibles, chacun composÃĐ de deux chromatides
soeurs reliÃĐes par un centromÃĻre. La membrane
nuclÃĐaire se dÃĐsagrÃĻge, permettant aux chromosomes
de s'aligner au centre de la cellule sous l'action
du fuseau mitotique, un rÃĐseau de microtubules
qui les guide. Les chromatides soeurs sont ensuite sÃĐparÃĐes et attirÃĐes
vers des pÃīles opposÃĐs de la cellule. Ce mouvement est assurÃĐ par le
raccourcissement des microtubules du fuseau.
Une fois les chromatides arrivÃĐes aux pÃīles, une nouvelle membrane nuclÃĐaire
se reforme autour de chaque groupe, reconstituant ainsi deux noyaux distincts.
ParallÃĻlement, le cytoplasme se divise, permettant la formation de deux
cellules filles.
à l'issue de la mitose, chaque cellule
fille possÃĻde le mÊme nombre de chromosomes que la cellule mÃĻre et est
prÊte à entrer dans un nouveau cycle cellulaire.
Ce processus est essentiel pour le maintien de l'intÃĐgritÃĐ gÃĐnÃĐtique
et joue un rÃīle fondamental dans la croissance et la rÃĐparation des tissus.
Les ÃĐtapes de la
mitose
La prophase.
Lors de la prophase
plusieurs ÃĐvÃĐnements clÃĐs se produisent pour prÃĐparer la cellule Ã
la division. L'aspect rÃĐticulÃĐ disparaÃŪt, et la chromatine
(ADN sous forme dÃĐcondensÃĐe) se trouve localisÃĐe en un long cordon pelotonnÃĐ
ou spirÃĻme. Ce cordon se raccourcit, se compacte, devient plus ÃĐpais
et moins pelotonnÃĐ, puis se coupe transversalement en plusieurs segments
appelÃĐs
chromosomes ou anses chromatiques.
Chaque chromosome est constituÃĐ de deux chromatides soeurs reliÃĐes par
un centromÃĻre. Le nuclÃĐole, qui est impliquÃĐ
dans la production des ribosomes, disparaÃŪt
progressivement.
Les centrosomes
(ou centrosphÃĻres chez les cellules animales) se dupliquent et migrent
vers les pÃīles opposÃĐs de la cellule, en organisant un rÃĐseau de microtubules
appelÃĐ fuseau mitotique. Ce dernier jouera un rÃīle essentiel dans la
sÃĐparation des chromatides soeurs. A la fin de la prophase ou au dÃĐbut
de la promÃĐtaphase, l'enveloppe nuclÃĐaire se dÃĐsagrÃĻge : la membrane
du noyau commence à se fragmenter, permettant aux microtubules du fuseau
mitotique d'interagir avec les chromosomes.
La promÃĐtaphase.
La promÃĐtaphase
une phase de transition oÃđ plusieurs ÃĐvÃĐnements se produisent pour assurer
la rÃĐpartition correcte des chromosomes. La disparition de la membrane
du noyau permet permet aux chromosomes d'interagir directement avec les
structures du fuseau mitotique. Chaque chromosome possÃĻde un centromÃĻre
contenant une structure spÃĐcialisÃĐe appelÃĐe kinÃĐtochore.
Les microtubules du fuseau mitotique se fixent aux kinÃĐtochores des chromatides
soeurs. Les chromosomes commencent à se dÃĐplacer activement, tirÃĐs et
poussÃĐs par les microtubules, jusqu'Ã s'aligner au centre de la cellule
en vue de la mÃĐtaphase. Cette phase assure que chaque chromosome est correctement
attachÃĐ au fuseau mitotique. Une connexion stable des chromosomes aux
microtubules est essentielle pour garantir une sÃĐparation correcte des
chromatides soeurs lors de l'anaphase.
La mÃĐtaphase.
La mÃĐtaphase est
caractÃĐrisÃĐe par l'alignement des chromosomes au centre de la cellule.
Les chromosomes, attachÃĐs aux microtubules du fuseau mitotique par leurs
kinÃĐtochores, sont dÃĐplacÃĐs et alignÃĐs au centre de la cellule, sur
un plan imaginaire appelÃĐ plaque ÃĐquatoriale. Un point de contrÃīle appelÃĐ
point de contrÃīle de la mÃĐtaphase (ou point de contrÃīle du fuseau) vÃĐrifie
que chaque chromatide soeur est bien attachÃĐe aux microtubules provenant
de pÃīles opposÃĐs. Cette vÃĐrification est nÃĐcessaire pour ÃĐviter des
erreurs de sÃĐparation des chromosomes, qui pourraient entraÃŪner des anomalies
gÃĐnÃĐtiques. Cette ÃĐtape garantit que la sÃĐparation des chromatides
soeurs se fera de maniÃĻre ÃĐquitable entre les deux cellules filles. Un
mauvais alignement ou une mauvaise fixation des chromosomes peut provoquer
des erreurs de distribution de l'ADN, ce qui peut mener à des anomalies
cellulaires.
-
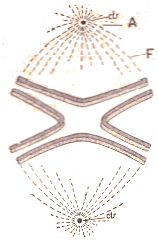 |
MÃĐtaphase.
Les quatre segments chromatiques, dÃĐdoublÃĐs chacun en deux chromosomes
sont rassemblÃĐs dans le plan ÃĐquatorial. A. Aster; F. Filaments achromatiques.
A chacun des pÃīles, il y a un centrosome entourÃĐ d'une vÃĐsicule directrice
et d'un aster (rayonnement de filaments achromatiques). La prÃĐsence d'un
aster à chaque pÃīle a valu à cette figure la dÃĐnomination d'amphiaster.
Les chromosomes sont orientÃĐs d'une maniÃĻre trÃĻs particuliÃĻre, chacun
d'eux avant la forme d'un U dont la connexitÃĐ est tournÃĐe vers le centre.
L'ensemble des chromosomes constitue la plaque ÃĐquatoriale (qui se compose
en rÃĐalitÃĐ de deux plaques parallÃĻles, chacune des plaques comprenant
la moitiÃĐ de chaque chromosome primitif dÃĐdoublÃĐ en deux anses jumelles
et parallÃĻles ombrÃĐes, l'une de hachures, l'autre de pointillÃĐ. |
L'anaphase.
L'anaphase correspond
à la sÃĐparation des chromatides soeurs et leur migration vers les pÃīles
opposÃĐs de la cellule. Une fois que le point de contrÃīle de la mÃĐtaphase
a validÃĐ l'attachement correct des chromosomes, une enzyme appelÃĐe sÃĐparase
coupe les liens entre les chromatides soeurs. Chaque chromatide devient
alors un chromosome indÃĐpendant. Les microtubules du fuseau mitotique
raccourcissent, tirant les chromosomes vers les pÃīles opposÃĐs de la cellule.
En parallÃĻle, d'autres microtubules s'allongent pour ÃĐloigner les pÃīles,
allongeant ainsi la cellule. Ã la fin de l'anaphase, chaque pÃīle de la
cellule possÃĻde une copie complÃĻte et identique du gÃĐnome
de la cellule mÃĻre. Lors de l'anaphase il est vÃĐrifiÃĐ que chaque cellule
fille recevra le mÊme nombre de chromosomes. Des erreurs dans cette phase
peuvent provoquer des anomalies chromosomiques, comme la non-disjonction,
qui peut entraÃŪner des maladies gÃĐnÃĐtiques (ex. trisomie).
La tÃĐlophase.
La tÃĐlophase marque
le retour à une organisation cellulaire normale avant la division complÃĻte
de la cellule. Une nouvelle membrane nuclÃĐaire se reforme autour des chromosomes
à chaque pÃīle de la cellule. Cela reconstitue deux noyaux distincts,
chacun contenant une copie complÃĻte du matÃĐriel gÃĐnÃĐtique. Les chromosomes,
qui ÃĐtaient trÃĻs condensÃĐs pendant la mitose, commencent à se dÃĐrouler
et à redevenir de la chromatine (forme plus relÃĒchÃĐe de l'ADN). Les
microtubules du fuseau mitotique se dÃĐsassemblent, car ils ne sont plus
nÃĐcessaires à la sÃĐparation des chromosomes. La cellule commence Ã
se diviser en deux par un processus appelÃĐ cytocinÃĻse,
qui va sÃĐparer complÃĻtement le cytoplasme et donner naissance à deux
cellules filles identiques. Cette phase permet de restaurer un ÃĐtat cellulaire
fonctionnel aprÃĻs la sÃĐparation des chromatides.Elle assure que chaque
cellule fille reçoit un noyau complet avant la division finale du cytoplasme.
Biochimie de la mitose
La mitose est un ÃĐvÃĐnement
hautement rÃĐgulÃĐ au niveau biochimique. Il existe une cascade d'ÃĐvÃĐnements
rÃĐgulÃĐs par des protÃĐines clÃĐs comme les
cyclines,
les CDK, les phosphatases,
les protÃĐines motrices, les condensines, les cohÃĐsines, les sÃĐparases
et l'APC/C. Ces acteurs biochimiques orchestrent la dynamique du fuseau
mitotique, la condensation et dÃĐcondensation des chromosomes, la rupture
et reformation de l'enveloppe nuclÃĐaire, la sÃĐparation prÃĐcise des chromatides
soeurs et la cytocinÃĻse, pour assurer la division cellulaire fidÃĻle et
la transmission du matÃĐriel gÃĐnÃĐtique aux cellules filles.
RÃĐgulation par
les cyclines et les kinases cycline-fÃĐpendantes (CDK).
Les cyclines et
les CDKs sont les moteurs biochimiques du cycle cellulaire. Les cyclines
sont des protÃĐines dont la concentration fluctue pendant le cycle
cellulaire, tandis que les CDK sont des kinases
(enzymes qui phosphorylent d'autres protÃĐines)
dont l'activitÃĐ dÃĐpend de leur liaison aux cyclines. Plusieurs complexes
cycline-CDK sont impliquÃĐs dans la mitose, les plus importants ÃĐtant
:
âĒ Cycline
B/CDK1 (MPF - Mitosis Promoting Factor). - Ce complexe est essentiel
pour l'entrÃĐe en mitose. Il dÃĐclenche de nombreux ÃĐvÃĐnements de la
prophase et promÃĐtaphase en phosphorylant de nombreuses protÃĐines cibles.
Son activitÃĐ augmente progressivement pendant l'interphase
et atteint un pic juste avant et pendant la mitose.
âĒ Cycline A/CDK2
et Cycline E/CDK2. - Bien que principalement impliquÃĐs dans les phases
S et G1/S, ils peuvent aussi avoir un rÃīle prÃĐparatoire à la mitose.
L'activitÃĐ des complexes
cycline-CDK est finement rÃĐgulÃĐe par la phosphorylation
et la dÃĐphosphorylation. Des kinases activatrices (CAK) et inhibitrices
(Wee1) phosphorylent la CDK1, et des phosphatases (CDC25) dÃĐphosphorylent
pour activer ou dÃĐsactiver le complexe cycline B/CDK1 au bon moment.
La dÃĐgradation des
cyclines par le systÃĻme ubiquitine-protÃĐasome est importante pour la
sortie de la mitose. L'APC/C (Anaphase Promoting Complex/Cyclosome)
est une ubiquitine ligase qui cible la cycline B pour la dÃĐgradation,
inactivant ainsi le CDK1 et permettant la sortie de la mitose.
Organisation et
dynamique du fuseau mitotique.
Le fuseau mitotique
est composÃĐ de microtubules, des polymÃĻres de tubuline.
La polymÃĐrisation (assemblage) et la dÃĐpolymÃĐrisation (dÃĐsassemblage)
dynamiques des microtubules sont essentielles pour la formation du fuseau,
la capture des chromosomes et leur mouvement. Ces processus sont ÃĐnergivores
et dÃĐpendent de GTP (guanosine triphosphate).
Les protÃĐines motrices,
comme les kinÃĐsines et les dynÃĐines, jouent un rÃīle dÃĐterminant dans
l'organisation du fuseau mitotique et
le mouvement des chromosomes. Elles utilisent l'hydrolyse de l'ATP (adÃĐnosine
triphosphate) pour se dÃĐplacer le long des microtubules et transporter
des charges, y compris les chromosomes.
Les centrosomes,
chez les animaux, ou les MTOC (microtubule
organizing center), chez les plantes et les
champignons,
sont les principaux centres organisateurs des microtubules. Leur duplication
et leur migration aux pÃīles opposÃĐs de la cellule sont des ÃĐvÃĐnements
charniÃĻres de la prophase.
Condensation et
dÃĐcondensation des chromosomes.
Les condensines
sont des complexes protÃĐiques impliquÃĐs dans la condensation des chromosomes
en structures compactes et organisÃĐes durant la prophase. Les condensines
s'associent à l'ADN et utilisent l'ÃĐnergie de l'ATP pour boucler et enrouler
les chromosomes, facilitant leur sÃĐgrÃĐgation.
Les topoisomÃĐrases
sont de enzymes essentielles pour dÃĐmÊler l'ADN
enchevÊtrÃĐ et permettre la condensation et la sÃĐparation des chromosomes
sans crÃĐer de noeuds ou de cassures.
Les histones,
protÃĐines autour desquelles l'ADN s'enroule, subissent des modifications
post-traductionnelles (phosphorylation, acÃĐtylation, mÃĐthylation) qui
influencent la structure de la chromatine et la condensation des chromosomes.
Rupture et reformation
de l'enveloppe nuclÃĐaire.
Les lamines sont
des protÃĐines fibreuses qui forment la lamina nuclÃĐaire, la structure
de soutien de l'enveloppe nuclÃĐaire. La phosphorylation
des lamines par la CDK1 en promÃĐtaphase provoque leur dÃĐpolymÃĐrisation
et la fragmentation de l'enveloppe nuclÃĐaire, permettant au fuseau mitotique
d'accÃĐder aux chromosomes.
Lors de la tÃĐlophase,
la dÃĐphosphorylation des lamines par des phosphatases (comme la phosphatase
PP1) conduit à leur repolymÃĐrisation et à la reformation de l'enveloppe
nuclÃĐaire autour de chaque lot de chromosomes fils.
SÃĐparation des
chromatides soeurs (anaphase).
Les chromatides
soeurs sont maintenues ensemble par des complexes protÃĐiques appelÃĐs
cohÃĐsines. La sÃĐparation des chromatides soeurs à l'anaphase est dÃĐclenchÃĐe
par la dÃĐgradation de la cohÃĐsine. La sÃĐparase, une protÃĐase, est activÃĐe
par l'APC/C et clive la sous-unitÃĐ Scc1 de la cohÃĐsine, permettant la
sÃĐparation des chromatides.
L'APC/C, complexe
ubiquitine ligase, joue un rÃīle important dans la transition mÃĐtaphase-anaphase.
Il cible la sÃĐcurine (un inhibiteur de la sÃĐparase) pour la dÃĐgradation,
activant indirectement la sÃĐparase et dÃĐclenchant la sÃĐparation des
chromatides soeurs. Comme mentionnÃĐ prÃĐcÃĐdemment, l'APC/C cible ÃĐgalement
la cycline B pour la dÃĐgradation, permettant la sortie de la mitose.
Points de contrÃīle
(checkpoints) mitotiques.
Le point de contrÃīle
du fuseau mitotique (spindle assembly checkpoint, SAC) surveille
l'attachement correct de tous les chromosomes au fuseau mitotique au niveau
des kinÃĐtochores avant de permettre la progression vers l'anaphase. Si
des chromosomes ne sont pas correctement attachÃĐs, le SAC active une voie
de signalisation qui inhibe
l'APC/C, bloquant ainsi la sÃĐparation des chromatides soeurs jusqu'Ã
ce que tous les attachements soient corrects. Le complexe MCC (mitotic
checkpoint complex) est un acteur clÃĐ de ce checkpoint.
Bien que moins directement
liÃĐs à la mitose elle-mÊme, des points de contrÃīle celui des dommages
à l'ADN (activÃĐ en G2) peuvent ÃĐgalement influencer
la progression vers la mitose en arrÊtant le cycle cellulaire si l'ADN
est endommagÃĐ.
CytocinÃĻse (division
du cytoplasme).
La cytocinÃĻse est
rÃĐalisÃĐe par un anneau contractile composÃĐ de filaments d'actine
et de myosine II. La contraction de cet anneau,
alimentÃĐe par l'ATP, ÃĐtrangle la cellule en deux cellules filles.
La formation et la
contraction de l'anneau contractile sont finement rÃĐgulÃĐes spatialement
et temporellement, dÃĐpendant de signaux provenant du fuseau mitotique
et de la rÃĐgion ÃĐquatoriale de la cellule.
La cytocinÃĻse implique
ÃĐgalement la partition des organites cellulaires
(mitochondries,
ribosomes,
etc.) entre les deux cellules filles, un processus qui peut Être alÃĐatoire
ou plus rÃĐgulÃĐ selon le type d'organite. |
|
