
ÂŦ
L'environnement n'est pas que co-prÃĐsent; il est aussi co-organisateur.
[...]. L'environnement, loin de rÃĐduire son caractÃĻre co-organisateur,
l'accroÃŪt chez l'Être vivant. Comme on le verra, l'environnement, devenu
ÃĐco-systÃĻme, c'est-Ã -dire une machine spontanÃĐe nÃĐe des interactions
entre les Êtres vivants d'une mÊme ÂŦ niche Âŧ, est beaucoup plus qu'une
rÃĐserve de nourriture, plus encore qu'une source de nÃĐguentropie oÃđ
l'Être puise de l'organisation, de la complexitÃĐ, de l'information, c'est
une des dimensions de la vie, aussi fondamentale que l'individualitÃĐ,
la sociÃĐtÃĐ, le cycle des reproductions.
Ainsi
s'impose l'idÃĐe clÃĐ : l'environnement est constitutif en permanence de
tous les Êtres qui s'alimentent en lui; il coopÃĻre en permanence avec
leur organisation. Ces Êtres et organisations sont donc en permanence
ÃĐco-dÃĐpendants. Mais, par un paradoxe qui est le propre de la relation
ÃĐcologique, c'est dans cette dÃĐpendance que se tisse et se constitue
l'autonomie de ces Êtres. De tels Êtres ne peuvent construire et maintenir
leur existence, leur autonomie, leur individualitÃĐ, leur originalitÃĐ
que dans la relation ÃĐcologique, c'est-Ã -dire dans et par la dÃĐpendance
à l'ÃĐgard de leur environnement ; d'oÃđ l'idÃĐe alpha de toute pensÃĐe
ÃĐcologisÃĐe : l'indÃĐpendance d'un Être vivant nÃĐcessite sa dÃĐpendance
à l'ÃĐgard de son environnement. Âŧ
Edgar
Morin
(La
MÃĐthode, I). |
On donne le nom
de biosphÃĻre à la partie de notre planÃĻte
oÃđ se rencontrent les organismes vivants. Cela
inclut la basse atmosphÃĻre, toute la rÃĐgion de la croÃŧte terrestre la
plus proche de la surface (la pÃĐdosphÃĻre, ÃĐpaisse de quelques mÃĻtres),
les ocÃĐans et les mers (hydrosphÃĻre), ainsi que l'ensemble des organismes
prÃĐsents dans cet espace.
On a proposÃĐ d'utiliser le terme d'ÃĐcosphÃĻre
pour dÃĐsigner la biosphÃĻre lorsqu'on envisage celle-ci non pas seulement
en tant que partie de la Terre abritant des organismes vivants, mais en
tant qu'ÃĐcosystÃĻme (ou ensemble de tous les ÃĐcosystÃĻmes), c'est-Ã -dire
lorsqu'on s'intÃĐresse à l'ensemble des processus en jeu lors des interactions
entre les diffÃĐrents organismes vivants et leur environnement. Cela implique
d'ÃĐtendre l'ÃĐtude à l'atmosphÃĻre
dans sa totalitÃĐ, puisque sa composition en oxygÃĻne (dioxygÃĻne O2
et ozone O3) dÃĐcoule entiÃĻrement de l'activitÃĐ
photosynthÃĐtique des plantes, et que les rejets dÃŧs aux activitÃĐs
humaines (gaz à effet de serre, CFC et autres polluants) ont ÃĐgalement
un impact global. Cela revient aussi à ajouter la dimension temporelle
et dynamique à l'ÃĐtude de cette biosphÃĻre et à l'envisager comme le
rÃĐsultat de processus à l'oeuvre depuis prÃĻs de quatre milliards d'annÃĐes,
c'est-Ã -dire depuis que les premiers organismes vivants sont apparus sur
la Terre.
L'ÃĐtude de la biosphÃĻre
et de l'ÃĐcosphÃĻre conduit, par la dÃĐfinition mÊme de son objet à considÃĐrer
deux ordres de phÃĐnomÃĻnes, ceux qui concernent
le vivant (facteurs biotiques) et ceux qui concernent tout ce qui n'est
pas vivant (facteurs abiotiques). Les interactions entre organismes vivants
(compÃĐtition, mutualisme, etc.), celles de caractÃĻre purement physique
et chimique (gÃĐophysique, gÃĐochimique), et celles entre les Êtres vivants
et leur milieu, qui dÃĐfinissent à proprement parler le domaine d'ÃĐtude
de l'ÃĐcologie. Ces interactions se font via des ÃĐchanges de matiÃĻre
et d'ÃĐnergie.
Les visages du vivant
L'ÃĐvolution du vivant.
La
chimie prÃĐbiotique.
L'existence d'organismes vivants
sur la Terre est attestÃĐe depuis , possiblement, 3,8 milliards d'annÃĐes
(soit seulement 800 millions d'annÃĐes aprÃĻs la formation de la planÃĻte).
Si l'on met à part l'ÃĐnergie indispensable
à toute la chimie du vivant, c'est la prÃĐsence d'eau liquide en abondance
qui a ÃĐtÃĐ la premiÃĻre condition à l'apparition de ces organismes. L'eau
liquide est un milieu qui permet à la fois la rencontre des ÃĐlÃĐments
chimiques qui peuvent se lier pour former des molÃĐcules de plus en plus
complexes. La molÃĐcule d'eau, est un agent rÃĐducteur (elle fournit un
ÃĐlectron lors de certaines rÃĐactions chimiques); en tant que dissolvant,
elle est en mesure de faciliter la rupture des liaisons entre molÃĐcules,
si bien qu'elle apparaÃŪt comme indispensable à toutes les transformations
chimiques opÃĐrÃĐes dans les organismes vivants.
L'autre ÃĐlÃĐment essentiel à la chimie
du vivant est le carbone. Les caractÃĐristiques de l'atome
de carbone (notamment sa tÃĐtravalence) le rendent apte à servir de ciment
aux ÃĐdifices molÃĐculaires ÃĐventuellement complexes (macromolÃĐcules)qui
sont à la base de tous les organismes vivants et des phÃĐnomÃĻnes qui
les caractÃĐrisent. Ces molÃĐcules sont formÃĐes d'un squelette d'atomes
de carbone auquel sont liÃĐs quantitÃĐ d'autres ÃĐlÃĐments (oxygÃĻne, hydrogÃĻne,
azote, calcium, phosphore, soufre, potassium, etc.). La chimie du carbone
est par excellence la chimie du vivant et est ordinairement nommÃĐe, pour
cette raison, chimie organique.
Les molÃĐcules organiques qui ont prÃĐcÃĐdÃĐ
les premiers organismes vivants se sont constituÃĐes à partir du carbone
disponible et de l'hydrogÃĻne, - celui-ci sans doute d'abord fourni par
les molÃĐcules de sulfure d'hydrogÃĻne H2S (agent
rÃĐducteur), puis par les molÃĐcules d'eau H2O. Certaines
de ces molÃĐcules (des acides aminÃĐs parmi elles) ont pu aussi Être synthÃĐtisÃĐes
dans l'espace et Être apportÃĐs sur la Terre par des chutes de
mÃĐtÃĐorites
ou des collisions avec des noyaux comÃĐtaires.
Le premier pas important sur le chemin qui menait aux organismes vivant
a ÃĐtÃĐ la formation des premiÃĻres molÃĐcules capables de se rÃĐpliquer.
Aujourd'hui, ce sont les molÃĐcules d'ADN qui, possÃĐdant cette capacitÃĐ,
portent l'information nÃĐcessaire (information gÃĐnÃĐtique) pour que puisse
se constituer un organisme vivant à partir d'une autre. Dans un premier
temps, ce rÃīle ÃĐtait dÃĐvolu à des molÃĐcules d'ARN ( Les
acides nuclÃĐiques), mais la grammaire gÃĐnÃĐrale de ce qui allait
Être la reproduction des organismes et de la constitution des espÃĻces
vivantes ÃĐtait la mÊme. Les
acides nuclÃĐiques), mais la grammaire gÃĐnÃĐrale de ce qui allait
Être la reproduction des organismes et de la constitution des espÃĻces
vivantes ÃĐtait la mÊme.
Descendance
et diffÃĐrence. DÃĻs l'ÃĐpoque prÃĐbiotique, lorsqu'on parle de molÃĐcules
capables de se dupliquer, on n'entend pas qu'elles produisent des rÃĐpliques
exactement identiques à elles-mÊmes. Il peut exister des diffÃĐrences
entre la molÃĐcule fille et la molÃĐcule mÃĻre, dues à des erreurs alÃĐatoires
dans la transmission de l'information gÃĐnÃĐtique. Certaines de ces erreurs
ont pour consÃĐquence une descendance plus ou moins apte à se reproduire
elle-mÊme (peut-Être en fonction du milieu dans lequel vivent les organismes
concernÃĐ, ou pour d'autres raisons). C'est le principe de l'ÃĐvolution,
qui va de pair avec la formation de divers chemins ÃĐvolutifs, certains
menant à l'apparition de nouvelles espÃĻces, d'autres à la disparition
d'espÃĻces existantes.
Un autre pas a ÃĐtÃĐ franchi quand des molÃĐcules
d'ARN capables de duplication se sont trouvÃĐes enfermÃĐes dans des microsphÃĻres.
De telles structures sont des membranes repliÃĐes sur elles-mÊmes (constituÃĐes
de
lipides et de protÃĐines),
qui ont pu se former spontanÃĐment, si les conditions physiques et mÃĐcaniques
ÃĐtaient rÃĐunies. Une molÃĐcule d'ARN capable de se rÃĐpliquer et enfermÃĐe
dans un espace dÃĐlimitÃĐ par une membrane, cela commence à ressembler
à une cellule vivante. Mais il a fallu encore
franchir quelques ÃĐtapes pour que l'on puisse parler d'organismes vivants.
Ces structures prÃĐbiotiques ont dÃŧ acquÃĐrir
notamment la capacitÃĐ de ce maintenir telles quelles malgrÃĐ les variations
possibles de leur environnement. Cela signifie, la capacitÃĐ de maintenir
des conditions physiques et chimiques à peu prÃĻs constantes à l'intÃĐrieur
de l'espace renfermÃĐ par la membrane. Cette capacitÃĐ d'autorÃĐgulation
interne qui, aprÃĻs toutes les caractÃĐristiques dÃĐjà ÃĐvoquÃĐes, forme
le socle de la dÃĐfinition d'un organisme vivant, est appelÃĐe
homÃĐostasie.
Un ÃĐlÃĐment central de cette capacitÃĐ de rÃĐgulation interne est la possibilitÃĐ
de gÃĐrer d'une maniÃĻre ou d'une autre l'ÃĐnergie qui provient (ou qui
ÃĐventuellement peut ne pas provenir pendant un certain temps) de l'extÃĐrieur.
Les toutes premiÃĻres cellules
vivantes, notamment grÃĒce à l'apparition chez elles, il y a environ
2,2 milliards d'annÃĐes, de certaines structures spÃĐcialisÃĐes, appelÃĐes
organites,
ont dÃŧ acquÃĐrir cette capacitÃĐ avant de perfectionner de diverses maniÃĻres
leurs modes d'acquisition de l'ÃĐnergie (comme la photosynthÃĻse apparue
trÃĻs prÃĐcocement), de transformation et de stockage interne. Ces processus,
en mÊme temps que ceux qui concernent la transformation de matiÃĻre Ã
l'intÃĐrieur d'un organisme vivant, en dÃĐfinissent le mÃĐtabolisme.
-

| Stromatolithes
de Shark Bay (Australie Occidentale). - Les stromatolithes sont des
formations minÃĐrales que l'on rencontre dans les zones intertidales (
= bandes littorales dÃĐlimitÃĐes par l'amplitude des marÃĐes). Elles sont
produites par des bactÃĐries, notamment par des cyanobactÃĐries. Ces micro-organismes
photosynthÃĐtiques dÃĐtachent le carbone du CO2 et
le libÃĻrent sous la forme de carbonates, qui, par prÃĐcipitation, forment
les stromatolithes. Ces formations ont existÃĐ Ã peu prÃĻs à tous les
moments de l'histoire de la Terre et se rencontrent sous des formes fossiles
en de nombreux lieux. Les stromatolithes ont connu leur ÃĒge d'or au ProtÃĐrozoÃŊque,
entre -2,2 milliards d'annÃĐes et -570 millions d'annÃĐes. Ils fournissent
certains des plus anciens indices de la prÃĐsence d'organismes vivants
sur notre planÃĻte. Des stromatolithes ont ainsi ÃĐtÃĐ dÃĐcouverts au Groenland,
vieux de 3,7 milliards d'annÃĐes. Photo : Paul Harrison
/ USGS. |
Procaryotes
et eucaryotes.
Dans les premiÃĻres cellules, les molÃĐcules
renfermant l'information gÃĐnÃĐtique ÃĐtaient simplement contenues à l'intÃĐrieur
de la membrane cellulaire, sans qu'un lieu bien dÃĐlimitÃĐ leur soit destinÃĐ,
comme c'est le cas encore aujourd'hui avec les bactÃĐries et les archÃĐes.
De tels organismes sont qualifiÃĐs de procaryotes. Mais l'ÃĐvolution
de certaines cellules les a conduites à enfermer les molÃĐcules renfermant
l'information gÃĐnÃĐtique dans une rÃĐgion spÃĐciale, elle aussi circonscrite
par une membrane, un organite particulier, appelÃĐe le noyau cellulaire.
Les cellules dotÃĐes d'un tel noyau (et les organismes formÃĐs de telles
cellules) sont dites eucaryotes.
Unicelluaires
et pluricellulaires.
Les Eucaryotes sont les organismes qui
ont suivi les chemins ÃĐvolutifs les plus variÃĐs. La plupart de ceux qui
sont restÃĐs constituÃĐs d'une seule cellule (organismes unicellulaires)
ont formÃĐ le gros du bataillon des Protistes dont les plus anciens remontent
à 1,8 milliards d'annÃĐes; les autres ont constituÃĐ des ensembles de
cellules fonctionnant ensemble pour former des organismes pluricellulaires.
Depuis une dÃĐcouverte faite au Gabon en 2008,
on fait remonter les plus anciens organismes pluricellulaires à 2,2 milliards
d'annÃĐes (Orosirien), mais ils pourraient avoir disparu assez rapidement.
Une autre souche, celle des algues rouges (Rhodophytes) remonte à 1, 2
milliards d'annÃĐes (StÃĐnien). Les ancÊtres des organismes pluricellulaires
actuels (les animaux, les vÃĐgÃĐtaux, les champignons) pourrait ne remonter
qu'Ã Ediacarien, il ya 600 millions d'annÃĐes (Le
PassÃĐ de la Terre). Quoi qu'il en soit, les organismes pluricellulaires
possÃĻdent des cellules spÃĐcialisÃĐes et en gÃĐnÃĐral des tissus diffÃĐrenciÃĐs,
chacun ayant ses propres caractÃĐristiques et fonctions, et qui forment
des organes.
Autotrophes
et hÃĐrÃĐrotrophes.
Tous les organismes
sont constituÃĐs deau, de sels minÃĐraux et de trÃĻs divers composÃĐs
organiques. Ils puisent dans le milieu oÃđ ils vivent les ressources (lumiÃĻre
du soleil, nutriments minÃĐraux et matiÃĻre organique) nÃĐcessaires Ã
leur survie, à leur croissance à leur entretien et à leur reproduction.
La nutrition fournit
les matÃĐriaux nÃĐcessaires à la construction des molÃĐcules dont l'organisme
est constituÃĐ. Celles-ci sont dÃĐgradÃĐes ou transformÃĐes de telle sorte
que des ÃĐchanges constants de matiÃĻre avec le milieu extÃĐrieur sont
nÃĐcessaires. L'ÃĐnergie, qu'elle soit apportÃĐe par la nutrition ou non,
est nÃĐcessaire dans la la plupart des processus mÃĐtaboliques, en
particulier ceux responsables de la construction des macromolÃĐcules Ã
partir de composÃĐs plus petits; elle aussi se dÃĐgrade en permanence et
doit Être renouvelÃĐe constamment.
Les organismes capables
de synthÃĐtiser leur propre nourriture (plus prÃĐcisÃĐment, capables d'utiliser
du carbone inorganique comme source de carbone), et de d'obtenir par eux-mÊme
l'ÃĐnergie dont ils ont besoin à partir de leur milieu sont appelÃĐes
autotrophes.
Ceux qui doivent se nourrir d'autres organismes pour acquÃĐrir leur ÃĐnergie
sont appelÃĐs hÃĐtÃĐrotrophes (en grec,
trophÃĻ = nourriture,
aliments).
âĒ
Les
autotrophes. - Un organisme autotrophe (au carbone) est un organisme
capable de synthÃĐtiser les molÃĐcules utilisÃĐes pour la construction
et le fonctionnement des Êtres vivants en puisant leur carbone dans des
composÃĐs simples (dioxyde de carbone CO2, dans le
cas des organismes terrestres, acide carbonique, H2CO3
pour les autotrophes marins). Dans certains cas l'acquisition du carbone
inorganique se fait à partir de rÃĐactions chimiques inorganiques (chimiosynthÃĻse),
mais le plus gÃĐnÃĐralement en utilisant l'ÃĐnergie lumineuse (photosynthÃĻse),
qui est ensuite stockÃĐe dans des liaisons covalentes qui relient les atomes
de carbone entre eux pour former notamment des glucides
(sucres).
+ Les photoautotrophes,
tels que les plantes, les algues et les bactÃĐries photosynthÃĐtiques,
exploitent l'ÃĐnergie solaire ( )
en la convertissant en ÃĐnergie chimique d'abord sous forme d'adÃĐnosine
triphosphate (ATP), et d'une enzyme qui untervient dans la photosynthÃĻse,
le nicotinamide adÃĐnine dinuclÃĐotide phosphate (NADP). L'ÃĐnergie stockÃĐe
dans l'ATP est ensuite utilisÃĐe pour synthÃĐtiser des molÃĐcules organiques
telles que le glucose (CH2O). Ainsi pourra-t-on
rÃĐsumer, au final, le processus de la photosynthÃĻse, par la rÃĐaction
: )
en la convertissant en ÃĐnergie chimique d'abord sous forme d'adÃĐnosine
triphosphate (ATP), et d'une enzyme qui untervient dans la photosynthÃĻse,
le nicotinamide adÃĐnine dinuclÃĐotide phosphate (NADP). L'ÃĐnergie stockÃĐe
dans l'ATP est ensuite utilisÃĐe pour synthÃĐtiser des molÃĐcules organiques
telles que le glucose (CH2O). Ainsi pourra-t-on
rÃĐsumer, au final, le processus de la photosynthÃĻse, par la rÃĐaction
:
CO2
+
H2O + 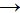 CH2O + O2
CH2O + O2
+ Les chimioautotrophes
sont principalement des bactÃĐries que l'on rencontre là oÃđ la lumiÃĻre
du soleil n'est pas disponible (grottes sombres, cheminÃĐes hydrothermales
au fond de l'ocÃĐan). De nombreux chimioautotrophes des cheminÃĐes hydrothermales
utilisent comme source d'ÃĐnergie chimique le sulfure d'hydrogÃĻne (H2S),
qu'elles libÃĻrent. Les chimioautotrophes, comme les photoautotrophes,
synthÃĐtisent ensuite des molÃĐcules organiques comme le glucose pour y
stocker l'ÃĐnergie requise par leur mÃĐtabolisme, et qui sera aussi utilisable
par d'autres organismes.
âĒ Les hÃĐtÃĐrotrophes.
- Un organisme hÃĐtÃĐrotrophe est un organisme incapable d'effectuer les
synthÃĻses de ses constituantsdirectement à partir d'ÃĐlÃĐments
minÃĐraux. Il doit puiser son ÃĐnergie et ses nutriments sous forme de
matiÃĻre organique en consommant d'autres organismes (autotrophes ou non).
En ce sens les hÃĐtÃĐrotrophes sont aussi chimiotrophes . Les animaux,
les champignons, certaines bactÃĐries, les protozoaires sont hÃĐtÃĐrotrophes.
Certains organismes,
rangÃĐs parmi les autotrophes, sont, Ã certains moments de leur vie, ou
pour certaines de leurs parties aussi hÃĐtÃĐrotrophes. On parle alors de
mixotrophie.
La plupart des Protistes sont mixotrophes. Il en est de mÊme des plantes
vasculaires (Cormophytes), qui sont sont autotrophes pour ce qui concerne
leur partie aÃĐrienne, mais hÃĐtÃĐrotrophes par leurs racines.
AnaÃĐrobies,
aÃĐrobies.
Les premiers organismes ÃĐtaient
anaÃĐorobies
: ils vivaient dans un environnement presque dÃĐpourvu d'oxygÃĻne (sa concentration
atmosphÃĐrique ÃĐtait infÃĐrieure à 2%). L'oxygÃĻne fabriquÃĐ par eux
(cyanobactÃĐries
ou leurs ancÊtres) au travers du processus de photosynthÃĻse, ÃĐtait aussitÃīt
fixÃĐ par divers ÃĐlÃĐments et minÃĐraux. Il ÃĐtait, par exemple, captÃĐ
par les
roches riches en fer (origine des formations
ferrifÃĻres rubanÃĐe du SidÃĐrien, caratÃĐrisÃĐes par la magnÃĐtite, Fe3O4).
Quand cette captation n'a plus ÃĐtÃĐ possible, faute de rÃĐcepteurs
disponibles (puits d'oxygÃĻne), cet ÃĐlÃĐment - toxique pour
les organismes anaÃĐrobies - s'est accumulÃĐ aussi bien en dissolution
dans l'eau que dans l'atmosphÃĻre. Beaucoup des organismes existants alors
ont disparu (on parle de catastrophe de l'oxygÃĻne pour qualifier cet ÃĐvÃĐnement
qui a eu lieu il y a 2,4 milliards d'annÃĐes).
Des CyanobactÃĐries ont cependant continuÃĐ
à enrichir l'atmosphÃĻre en oxygÃĻne. Il a fallu du temps pour que se
dÃĐveloppent des organismes aÃĐrobies, c'est-Ã -dire capables non
seulement de vivre dans un milieu riche en oxygÃĻne, mais d'utiliser cet
oxygÃĻne dÃĐsormais disponible en abondance. L'utilisation de l'oxygÃĻne
par les processus mÃĐtaboliques correspond à la respiration.
La
respiration aÃĐrobie se prÃĐsente ainsi comme le processus inverse de la
photosynthÃĻse.
On peut en schÃĐmatiser les effets chimiques par
la rÃĐaction :
CH2O
+ O2
CO2 + H2O +
(La combustion lente
du glucose libÃĻre l'ÃĐnergie renfermÃĐe dans les liaisons molÃĐculaires
de celui-ci et produit du dioxyde de carbone et de l'eau).
La concentration de
l'oxygÃĻne dans l'atmosphÃĻre a crÃŧ Ã peu prÃĻs rÃĐguliÃĻrement, pour
s'ÃĐtablir, il y a environ 500 milions d'annÃĐes à des niveaux du mÊme
ordre que ceux que l'on constate aujourd'hui. Chemin faisant l'oxygÃĻne
atmosphÃĐrique a permis un refroidissement du climat terrestre en faisant
diminuer la concentration atmosphÃĐrique du mÃĐthane (un gaz à effet de
serre).
A la fin du ProtÃĐrozoÃŊque,
l'oxygÃĻne, sous la forme d'ozone (trioxygÃĻne) a constituÃĐ dans la haute
atmosphÃĻre une couche capable de bloquer les rayonnements ultraviolets
du Soleil les plus nocifs pour les organismes vivants. Ceux-ci ÃĐtaient
jusque là confinÃĐs dans les ocÃĐans ou du moins dans des milieux aquatiques;
l'augmentation de l'oxygÃĻne et la diminution du risque ultraviolet ont
facilitÃĐ leur ÃĐtablissement sur la terre ferme et donnÃĐ un grand ÃĐlan
au dÃĐveloppement des organismes pluricellulaire terrestres, d'abord vÃĐgÃĐtaux
puis animaux. Ces conditions nouvelles ont ainsi prÃĐparÃĐ la grande ÃĐpoque
du vivant, marquÃĐe par une diversitÃĐ de formes inÃĐdites, connue sous
le nom d'explosion cambrienne (vers 540-530 millions d'annÃĐes avant le
prÃĐsent). ParallÃĻlement, le phytoplancton a aussi connu un essor considÃĐrable.
Il rÃĐalise, Ã l'ÃĐpoque actuelle, 40 % de toute la photosynthÃĻse.
-
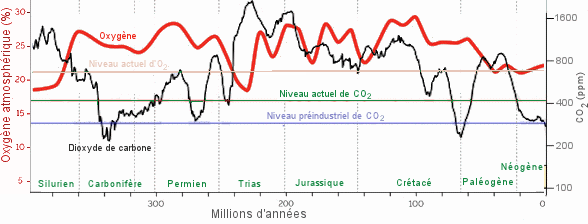
| Evolution
de la concentration de l'oxygÃĻne (rouge) et du dioxyde de carbone
(noir) dans l'atmosphÃĻre pendant le PhanÃĐrozoÃŊque (du Cambrien à aujourd'hui).
- L'atmosphÃĻre est restÃĐe rÃĐductrice jusqu'Ã il y a environ 600 milions
d'annÃĐes. A partir de là , la concentration d'oxygÃĻne, aprÃĻs plusieurs
oscillations, avec un pic autour de 20% au moment de l'explosion du Cambrien,
est passÃĐe de 10% Ã un niveau proche de celui qu'elle a actuellement,
soit environ 21%. Au cours des derniers 400 millions d'annÃĐes, elle n'a
oscillÃĐ que dans des limites relativement ÃĐtroites autour de 24%. Ce
sont les organismes photosynthÃĐtiques qui en sont responsables. Le CarbonifÃĻre,
le Permien et le CrÃĐtacÃĐ
prÃĐsentaient des concentrations d'oxygÃĻne ÃĐlevÃĐes; le Trias
infÃĐrieur et moyen a ÃĐtÃĐ une pÃĐriode de plus faible teneur en oxygÃĻne;
du milieu du Trias supÃĐrieur au
Jurassique,
l'oxygÃĻne semble avoir fluctuÃĐ mais est restÃĐ supÃĐrieur aux niveau
actuel. Depuis le CrÃĐtacÃĐ, le niveau d'oxygÃĻne a diminuÃĐ.
Il
y a beaucoup moins de CO2 dans
l'atmosphÃĻre que d'oxygÃĻne, et les variations de la concentration de
dioxyde de carbone sont reprÃĐsentÃĐes sur le graphique à une ÃĐchelle
trÃĻs diffÃĐrente (en ppm = parties par million). On notera la grande chute
de la concentration de CO2 pendant
le CarbonifÃĻre, c'est-à -dire à l'ÃĒge d'or des cryptogames
vasculaires (des ptÃĐridophytes, notamment).
Le carbone fixÃĐ ainsi par ces vÃĐgÃĐtaux est l'actuel rÃĐservoir des combustibles
fossiles. Comme celle de l'oxygÃĻne, la concentration du dioxyde de carbone
a beaucoup oscillÃĐ. Le lissage de la courbe ne permet pas de figurer des
variations de courte durÃĐe, tels les pics à la fin du Permien ou du CrÃĐtacÃĐ
ou encore la montÃĐe vertigineuse du CO2 atmosphÃĐrique
(de 278 ppm à 420 ppm) depuis le dÃĐbut de l'ÃĻre industrielle.
(Sources
: K. Cantner, AGI et Foster et al.). |
Le vivant en ses
royaumes.
Dans les classifications
habituellement utilisÃĐes du monde vivant, on
distingue six rÃĻgnes (archÃĐes, bactÃĐries, protistes, vÃĐgÃĐtaux, champignons
et animaux), entre lesquels se distribuent toutes les espÃĻces actuelles.
Les
ArchÃĐes et les BactÃĐries.
Les archÃĐes et les bactÃĐries
sont des organismes procaryotes et presque tous unicellulaires. Ils sont
presque tous microscopiques (les plus grosses bactÃĐries peuvent cependant
atteindre presque un millimÃĻtre de longueur). Ces deux groupes, dont l'ÃĐvolution
sÃĐparÃĐe a commencÃĐ Ã peu prÃĻs 200 millions d'annÃĐes aprÃĻs l'apparition
des premiers organismes vivants, se distinguent par leur biochimie. On
les rencontre principalement dans les zones humides.
Les
Protistes.
Les protistes sont gÃĐnÃĐralement des
organismes unicellulaires. Certains, Ã l'image du phytoplancton qui dÃĐrive
dans les ocÃĐans et les
lacs,
sont photosynthÃĐtiques, comme les plantes et
utilisent donc l'ÃĐnergie du soleil pour fabriquer de la nourriture. D'autres,
à l'image des amibes, du blob (Physarum polycephalum) ou des paramÃĐcies,
ont davantage d'affinitÃĐs avec les animaux, on les range parmi les protozoaires.
Les protistes vivent dans l'eau ou dans les milieux humides.
Les
VÃĐgÃĐtaux.
Les vÃĐgÃĐtaux
sont des organismes pluricellulaires vivant sur la terre et parfois dans
l'eau. Certains sont trÃĻs simples (mousses,
fougÃĻres,
ou les algues, ces derniÃĻres parfois rangÃĐes
parmi les protistes), d'autres plus complexes, Ã l'image des spermatophytes
(plantes à graines). Tous ont pour caractÃĐristique de pouvoir utiliser
presque toute l'ÃĐnergie qu'ils reçoivent de la lumiÃĻre solaire pour
opÃĐrer la photosynthÃĻse.
Les premiÃĻres algues
marines remontent au Cambrien. Les premiÃĻres plantes vasculaires
apparaissent à l'Ordovicien, elle commencent à envahir la terre, pour
prendre une grande extension au CarbonifÃĻre (grandes forÊts de fougÃĻres
arborescentes et de lycopodes se dÃĐveloppent,
premiers
conifÃĻres), qui correspond aussi
à l'apparition des premiers pollens. Les premiÃĻres
forÊts de gymnospermes (conifÃĻres, etc.)
remontent au Triassique. Les plantes à graines deviennent dominantes Ã
partir du CrÃĐtacÃĐ.
Les
Champignons.
Les champignons,
comme les vÃĐgÃĐtaux parmi lequels on les rangeait autrefois, sont gÃĐnÃĐralement
fixÃĐs au sol ou sur un support solide, et possÃĻdent des racines
(leur systÃĻme racinaire est souvent bien plus vaste que leur partie visible,
et peut atteindre plusieurs kilomÃĻtres carrÃĐs pour certaines espÃĻces).
Mais, dÃĐpourvus de la capacitÃĐ de photosynthÃĻse, il sont rangÃĐs aujourd'hui
dans un rÃĻgne distinct. Ils se nourrisent de matiÃĻre organique (de plantes
mortes, en particulier). Les champignons sont gÃĐnÃĐralement pluricellulaires,
mais quelques-uns, les levures (saccharomycÃĻtes), sont microscopiques
et unicellulaires. Les plus anciens champignons remontent au Silurien
ou à la fin de l'Ordovicien. (Certains champignons
infÃĐrieurs, gÃĐnÃĐralement photosynthÃĐtiques, peuvent aussi Être dÃĐtachÃĐs
des champignons proprement dits, et constituer un rÃĻgne supplÃĐmentaire,
celui des chromistes).
Les
Animaux.
Certains animaux
ne mesurent pas plus d'un millimÃĻtre de longueur, d'autres, comme la Baleine
bleue (Balaenoptera musculus), peuvent atteindre les 30 m. Les animaux,
qui vivent aussi bien sur la terre que dans les milieux aquatiques, se
distinguent des plantes par leur incapacitÃĐ Ã synthÃĐtiser leurs aliments
(Ã commencer par les sucres) Ã partir de composÃĐs chimiques bruts. Ils
survivent en mangeant d'autres Êtres vivants, ou des choses qui ÃĐtaient
autrefois vivantes (des plantes et des champignons, voire des protistes
(plancton), ou d'autres animaux). Si l'on excepte
queques animaux aquatiques qui restent pratiquement toute leur vie arrimÃĐs
à un support fixe (les mouvements de l'eau leur apportant la nourriture
nÃĐcessaire), la plupart des animaux sont capables de locomotion, soit
grÃĒce à des organes spÃĐcialisÃĐs (nageoires,
ailes,
pattes),
soit par les mouvements de leur corps tout entier. La plupart des animaux
ont une
tÊte sur laquelle sont placÃĐs plusieurs
organes sensoriels (yeux, narines, oreilles),
qui fournissent une aide pour les dÃĐplacements et l'identification de
la nourriture. Il est commode de distinguer parmi les animaux deux groupes
:
+ Les InvertÃĐbrÃĐs, qui
reprÃĐsentent plus de 95 % des espÃĻces animales actuellement connues.
Ils ne possÃĻdent pas de colonne vertÃĐbrale et
de squelette interne articulÃĐ. Parmi eux : les vers, les insectes,
les mollusques,
les cnidaires (mÃĐduses, coraux), etc.
+ Les VertÃĐbrÃĐs,
trÃĻs minoritaires, qui possÃĻdent, comme leur nom l'indique, une colonne
vertÃĐbrale et un squelette interne. Parmi
eux : les poissons, les amphibiens,
les reptiles, les oiseaux
et les mammifÃĻres.
Les premiers animaux
ÃĐtaient des invertÃĐbrÃĐs marins au corps mou, et sont apparus il y a
environ un milliard d'annÃĐes. L'explosion cambrienne a profitÃĐ Ã l'essor
des animaux (mÃĐtazoaires) : spongiaires,
arthropodes,
vertÃĐbrÃĐs. La plupart des invertÃĐbrÃĐs marins , Ã commencer par les
cÃĐphalopodes,
remontent à l'Ordovicien. A cette ÃĐpoque, les arthropodes, les cÃĐphalopodes
et les autres mollusques commencent à dominer
les mers, et sont rejoints par les poissons, de plus en plus prÃĐsents
à partir du Silurien. Au DÃĐvonien, les insectes
et les premiers tÃĐtrapodes apparaissent. Les amphibiens, les reptiles
et les insectes formeront les principales classes d'animaux jusqu'Ã l'apparition
des dinosaures (Triassique) et des mammifÃĻres
(Jurassique). La disparition des Dinosaures lors de la transition CrÃĐtacÃĐ-Tertiaire,
il y a 65 millions d'annÃĐes, permettra aux MammifÃĻres de prendre une
place de plus en plus importante.
Nombre d'espÃĻces
rÃĐpertoriÃĐes et nombre estimÃĐ d'espÃĻces
| Domaine |
RÃĻgne |
EspÃĻces
rÃĐpertoriÃĐes |
Nombre
d'espÃĻces
estimÃĐ |
| Procaryotes |
ArchÃĐes |
175 |
? |
| BactÃĐries |
10
000 |
? |
| Eucaryotes |
Protistes |
80
000 |
600
000 |
| Champignons |
72
000 |
1
500 000 |
| Plantes |
270
000 |
320
000 |
| Animaux |
|
| Arthropodes |
| |
Mandibulates
CrustacÃĐs
ChÃĐlicÃĐrates |
Mollusques
NÃĐmatodes
| VertÃĐbrÃĐs |
| |
Poissons
Amphibiens
Reptiles
Oiseaux
MammifÃĻres |
|
|
| - |
|
70 000
25
000
| 52
500 |
| |
25
000
4950
8002
9750
4630 |
|
|
| 1
320 000 |
|
| - |
|
8
000 000
150 0000
750
000 |
200 000
400
000
|
|
| Total |
1
750 000 |
14
000 000 |
Sources
: UNEP / World Atlas of biodiversity, 2003.
Les grandes divisions
de la biosphÃĻre et de l'ÃĐcosphÃĻre
Les organismes vivants
se rÃĐpartissent dans la biosphÃĻre en fonction des conditions qui leur
sont offertes, notamment selon la disponibilitÃĐ de l'eau, de l'ÃĐnergie
et de la nourriture.
Les
ÃĐcozones.
Les ÃĐcozones
sont les plus grandes divisions biogÃĐographiques de la surface de la Terre,
et se distinguent les unes des autres par des distributions caractÃĐristiques
de la flore et des animaux qui y vivent. Elles correspondent à des rÃĐgions
oÃđ les plantes et les animaux se sont dÃĐveloppÃĐs dans un isolement relatif
pendant de longues pÃĐriodes, du fait de divers facteurs gÃĐographiques
(ocÃĐans, chaÃŪnes de montagnes,
dÃĐserts,
etc), qui ont constituÃĐ des obstacles à la migration des plantes ou des
animaux. Une ÃĐcozone est donc dÃĐfinie selon les relations de parentÃĐ
entre les espÃĻces vivantes qu'on y rencontre. Ces relations atant elles-mÊmes
corrÃĐlÃĐes avec une histoire gÃĐographique et gÃĐologique commune. On
en compte huit.
âĒ
La
rÃĐgion palÃĐarctique comprend l'Europe, l'Afrique
septentrionale, le nord et le centre de la PÃĐninsule
arabique, ainsi que l'Asie au nord de l'Himalaya.
On y range l'Islande, les ÃŪles de l'Arctique
eurasiatique et l'archipel japonais (sauf l'archipel
Ryukyu). Cela reprÃĐsente prÃĻs de 55 millions de kilomÃĻtres carrÃĐs.
âĒ La rÃĐgion
nÃĐarctique comprend la plus grande partie de l'AmÃĐrique
du Nord (dont le Groenland), au jusqu'au
Sud du Mexique (Hauts plateaux).
âĒ La rÃĐgion
nÃĐotropicale, commence au Sud du Mexique (Terres chaudes) et s'ÃĐtend
jusqu'au Sud de l'AmÃĐrique du Sud (rÃĐgions
tropicales et rÃĐgions tempÃĐrÃĐes). Elle comprend aussi les Antilles.
âĒ La rÃĐgion
afrotropicale ou ÃĐthiopique comprend toute l'Afrique
subsaharienne, Madagascar et les ÃŪles voisinnes
le sud de la PÃĐninsule arabique, ainsi que le Sud de l'Iran
et du Pakistan.
âĒ La rÃĐgion
indomalaise ou orientale s'ÃĐtend sur la plus grande partie
de l'Asie du Sud, du Sud du Pakistan au Sud de la Chine
(provinces du Guangxi, Yunnan et Hainan) et jusqu'aux Philippines,
à Taiwan, aux ÃŪles Ryukyu , Ã
BornÃĐo
et à l'archipel indonÃĐsien (à l'Ouest de
Lombok).
âĒ La rÃĐgion
australasienne recouvre à peu prÃĻs ce que l'on appelle l'Australasie.
Elle est sÃĐparÃĐe de la rÃĐgion indo-malaise par la ligne Wallace (entre
Bali
et Lombok). Elle comprend l'est de l'IndonÃĐsie,
l'Australie, la Nouvelle-GuinÃĐe.
On y ajoute parfois plusieurs groupes d'ÃŪles du Pacifique,
que l'on dÃĐtache de la rÃĐgion ocÃĐanienne : le Vanuatu,
les ÃŪles salomon, la Nouvelle-ZÃĐlande,
La Nouvelle CalÃĐdonie, etc.
âĒ La rÃĐgion
ocÃĐanienne comprend les ÃŪles du Pacifique de PolynÃĐsie,
de MicronÃĐsie et de MÃĐlanÃĐsie
(sauf ÃĐventuellement les ÃŪles prÃĐcÃĐdemment nommÃĐes).
âĒ La rÃĐgion
antarctique comprend l'Antarctide et
divers groupes du Sud de l'ocÃĐan Indien
(ÃŪles Crozet, Kerguelen,
etc) et de l'ocÃĐan Atlantique (ÃŪles
Shetland du Sud et GÃĐorgie du Sud,
etc.).
Les limites entre les
ÃĐcozones ne sont pas toujours bien dÃĐfinies. A l'image de la ligne Wallace
(L'OcÃĐanie),
ces limites sont moins des lignes que des bandes plus ou moins ÃĐtendues,
qui dÃĐfinissent des ÃĐcoclines, autrement dit des gradations entre
deux zones. A l'intÃĐrieur de ces zones de transition entre deux faunes
et flores caractÃĐristiques des ÃĐcozones concernÃĐes, diffÃĐrentes lignes
pouvent souvent Être tracÃĐes en fonction des groupes de plantes ou d'animaux
considÃĐrÃĐs.
Les
ÃĐcorÃĐgions.
Les ÃĐcorÃĐgions
sont des zones gÃĐographiques ÃĐtendues auxquelles les divers caractÃĻres
(climat, gÃĐomorphologie, hydrologie, faune et flore, ÃĐventuellement les
caractÃĻres rÃĐunis confÃĐrÃĐs par l'activitÃĐ humaine) confÃĻrent une
ÃĐcologie particuliÃĻre. L'Amazonie, les Everglades
en Floride, la forÊt du Sri
Lanka, la savane soudanienne, la forÊt mixte de l'Europe
mÃĐditerranÃĐenne, par exemple, sont des ÃĐcorÃĐgions.
Les
biomes.
La notion de biome
se rapproche de celle d'ÃĐcorÃĐgion, mais possÃĻde un caractÃĻre plus gÃĐnÃĐrique
(ex. : la taÃŊga sibÃĐrienne et la taÃŊga canadienne sont des ÃĐcorÃĐgions;
la taÃŊga tout court est un biome). Les biomes sont des zones de la biosphÃĻre
dÃĐfinies par le climat et la gÃĐographie et prÃĐsentant des conditions
climatiques similaires. Ils sont dÃĐfinis sur la base de facteurs tels
que les structures vÃĐgÃĐtales (arbres, arbustes et herbes), les types
de feuilles (feuilles larges et aiguilles), l'espacement des plantes (forÊt,
bois, savane) et le climat. Contrairement aux ÃĐcorÃĐgions et aux ÃĐcozones,
les biomes ne sont pas dÃĐfinis par des similitudes gÃĐnÃĐtiques, taxonomiques
ou historiques.
On distingue les
biomes
terrestres (rÃĐpartis en trois groupes principaux : forÊts, formations
herbacÃĐes, dÃĐserts), les biomes aquatiques et les biomes mixtes.
Ces-derniers sont dÃĐfinis à partir des conditions particuliÃĻres nÃĐes
de la rencontre de zones terrestres et de zones aquatiques (eaux peu profondes,
plages, bords de riviÃĻres, etc.) .
Les biomes terrestres
reçoivent leur nom en fonction de leur formation vÃĐgÃĐtale typique et
pleinement dÃĐveloppÃĐe ou d'une caractÃĐristique marquante de son environnement
abiotique. Les caractÃĐristiques du macroclimat existant sont souvent utilisÃĐes
à cet effet. Les principaux biomes terrestres sont signalÃĐs en gras dans
le tableau ci-dessous. Ajoutons, que les biomes terrestres peuvent Être
divisÃĐes en entitÃĐs plus petites, les paysages, possÃĐdant une
physionomie propre.
L'oecumÃĻne.
- Ce mot dÃĐsigne l'ensemble des terres habitÃĐes en permanence et transformÃĐes
directement par l'espÃĻce humaine. Elle est formÃĐe d'un ensemble de biomes,
dits anthropogÃĐniques, dans lesquels on distingue divers types de paysages
(paysages urbains, paysages ruraux, etc.). On estime que l'oecumÃĻne recouvre
aujourd'hui la moitiÃĐ de la surface terrestre.
Les biomes aquatiques
sont le plus souvent ÃĐtÃĐ dÃĐfinis à partir de caractÃĻres purement gÃĐographiques;
division en biomes continentaux (lacs, cours d'eau) et en biomes ocÃĐaniques,
pour lesquels (comme dans le cas de certains lacs) la notion de profondeur
est prise en compte, car elle commande notamment à l'accÃĻs à l'ÃĐnergie
solaire. Les rÃĐgions supÃĐrieures sont atteintes par la lumiÃĻre (on parle
de zone photique) et permettent la photosynthÃĻse. Dans les rÃĐgions
infÃĐrieures, sombres, (zone aphotique), l'accÃĻs à l'ÃĐnergie (cela signifie
aussi à la nourriture) doit se faire selon des modes particuliers.
-
Les principaux
biomes
| ForÊts |
ForÊts subarctiques |
ForÊts de conifÃĻres
(taÃŊga, en Russie) |
| ForÊts
tempÃĐrÃĐes |
ForÊts tempÃĐrÃĐes
de conifÃĻres tempÃĐrÃĐes |
| ForÊts humides
(feuillus ou conifÃĻres) |
| ForÊts de feuillus |
| ForÊts mixtes (espÃĻces
dÃĐcidues et sempervirentes mÃĐlangÃĐes) : forÊts mÃĐditerranÃĐennes
ou sclÃĐrophylles
(bois, broussailles, maquis); forÊts de feuillus
et de conifÃĻres |
ForÊts
tropicales
et subtropicales |
ForÊts pluviales
ÃĐquatoriales |
| ForÊts de
mousson
(sÃĻches par pÃĐriodes) |
| ForÊts tropophiles
(forÊts tropicales sÃĻches) |
| FÃīrets tropicales
de conifÃĻres (forÊts de pins, forÊts de cyprÃĻs). |
| Formations
herbacÃĐes |
Savanes |
Savanes herbeuses
(Llanos, en AmÃĐrique du Sud). |
| Savanes arbustives |
| Savanes ÃĐpineuses,
broussailles |
| Landes,
bruyÃĻres |
| Prairies |
| Steppes |
Steppes proprement
dites, pampas, veldt, etc. selon les rÃĐgions |
| Toundras |
Toundras arctiques |
| Toundras alpines
(hauts plateaux) |
| DÃĐserts |
DÃĐserts
froids |
Glaces : glaciers,
inlandsis (des
microorganismes ont ÃĐtÃĐ dÃĐcouverts en 2014, vivant à 800 m de profondeur
sous les glaces de l'Antarctide). |
| Rochers |
| DÃĐserts
chauds |
DÃĐsert de sable
(erg, au Sahara) |
| DÃĐsert de roches
(reg, hammada, sÃĐrir, au Sahara) |
| Oasis |
| Biomes
anthropogÃĐniques |
Terres agricoles |
Terres labourÃĐes,
jardins, vergers, vignobles, riziÃĻres, pÃĒturages,
etc. |
| Zones urbanisÃĐes |
Villes, habitats
humains divers, bordures des voies de communication (routes, voies ferrÃĐes). |
Biomes
aquatiques
(hydrobiosphÃĻre) |
Eaux
continentales
(environ
1,8 % de la surface du globe) |
Lacs,
ÃĐtangs |
| Cours
d'eau : ruisseaux, riviÃĻres, fleuves |
| Deltas |
Eaux
ocÃĐaniques
(environ
70 % de la surface du globe) |
OcÃĐans ouverts
(milieu pÃĐlagique, prÃĻs de la surface; milieu dÃĐmersal
à des profondeurs plus grandes) |
| Grandes profondeurs,
fosses abyssales |
| Plateaux continentaux,
mers fermÃĐes, zones nÃĐritiques ou sublittorales |
| Zones d'algues flottantes |
| Banquises |
| Biomes
mixtes |
Milieux
littoraux |
Zones intertidales
(espaces entre les marÃĐes hautes et basses)
: vasiÃĻres, plages, estuaires |
| Mangroves |
| Zones
ripariennes (bordures de riviÃĻres et de lacs), forÊts-galeries |
| Zones
humides : Marais, prairies, savanes et forÊts
inondÃĐes ou inondables |
| RÃĐcifs
coralliens (atolls) |
| Zones
benthiques |
Fonds ocÃĐaniques,
fonds lacustres; limons |
| ForÊts d'algues,
prairies sous-marines |
| Events ocÃĐaniques
: cheminÃĐes hydrothermales, ÃĐvents froids, sources sous-marines |
| Montagnes
: ÃĐtagement de plusieurs biomes selon l'altitude.
Une grande variÃĐtÃĐ d'ÃĐcosystÃĻmes, reprÃĐsentatifs de diffÃĐrents biomes
peuvent ainsi se rencontrer dans des espaces trÃĻs rÃĐduits. |
Biomes des
milieux extrÊmes |
Milieux
dans lesquels les conditions physiques et chimiques "extrÊmes" : sources
hydrothermales, pergÃĐlisol, grottes, l'intÃĐrieur des roches et
des coraux, centrales nuclÃĐaires, etc. |
Les ÃĐcosystÃĻmes
Les systÃĻmes dynamiques.
Au sens le plus
large, un systÃĻme est un ensemble dont les ÃĐlÃĐments entretiennent entre
eux des relations qui les rendent interdÃĐpendants et confÃĻrent ainsi
une structure à cet ensemble. Cette dÃĐfinition conduit à s'interroger
sur ce que devient le systÃĻme au fil du temps, du fait mÊme de l'existence
de telles relations.
Lorsque ces relations
induisent des modifications dans le systÃĻme on parle d'interactions. Un
systÃĻme concret ÃĐvoluant ainsi en fonction des interactions entre ces
ÃĐlÃĐments est appelÃĐ systÃĻme dynamique. Un tel systÃĻme peut
conserver ses caractÃĐristiques (les paramÃĻtres qui le dÃĐfinissent)
dans la durÃĐe, se transformer pour se placer dans un ÃĐtat diffÃĐrent,
ou, Ã l'inverse, perdre complÃĻtement sa structure pour se disloquer.
La grandeur physique
utilisÃĐe pour caractÃĐriser une transformation et sa possibilitÃĐ Ã l'intÃĐrieur
d'un systÃĻme est l'ÃĐnergie : on envisage les trasformations à l'intÃĐrieur
d'un systÃĻme par le biais des ÃĐchanges d'ÃĐnergie entre les diffÃĐrentes
composantes du systÃĻme lors de leurs interactions. Dans un systÃĻme fermÃĐ,
l'ÃĐnergie (et la matiÃĻre) dont use le systÃĻme est limitÃĐe par la dÃĐfinition
mÊme du systÃĻme. Au fil des transformations, elle perd de son efficacitÃĐ
(deuxiÃĻme principe de la thermodynamique) et, au final, aucune modification
à l'intÃĐrieur du systÃĻme n'est plus possible. A l'inverse, dans le cas
d'un systÃĻme ouvert, celui-ci est alimentÃĐ de l'extÃĐrieur en
ÃĐnergie (et ÃĐventuellement en matiÃĻre). L'ÃĐvolution du systÃĻme se
comprend alors en termes de flux d'ÃĐnergie : l'ÃĐnergie entrante permet
les transformations à l'intÃĐrieur du systÃĻme, et l'inÃĐvitable dÃĐgradation
de l'ÃĐnergie (qui la rend de moins en moins efficace) est compensÃĐe par
le maintien du flux entrant. Lorsque, sous l'effet d'un flux entrant rÃĐgulier
un systÃĻme est en mesure d'actionner ses diffÃĐrents composants afin de
maintenir son ÃĐquilibre, il est dit dans un ÃĐtat stationnaire.
Quand on raisonne
en termes de flux, deux types de processus, appelÃĐs boucles de rÃĐtroaction,
interviennent dans l'ÃĐvolution, l'ÃĐquilibre ou non, d'un systÃĻme, c'est-Ã -dire
dans sa rÃĐgulation :
âĒ
La rÃĐtroaction positive correspond à la situation oÃđ les effets
d'un processus agissent de telle sorte que ce processus soit renforcÃĐ
(effet boule de neige). Ce type de rÃĐtroaction est nÃĐcessaire pour activer
ou stimuler une transformation, une interaction, mais il aboutirait Ã
un emballement incontrolable si un deuxiÃĻme type de processus n'intervenait
pas :
âĒ La rÃĐtroaction
nÃĐgative correspond à la situation oÃđ les effets d'un processus
font que le moteur de celui-ci est inhibÃĐ (effet de soupape).
Ces deux processus combinÃĐs
dÃĐfinissent la maniÃĻre dont le systÃĻme se rÃĐgule. La capacitÃĐ d'un
systÃĻme à se rÃĐguler de telle sorte que ces conditions internes restent
confinÃĐes dans des limites gÃĐnÃĐralement ÃĐtroites est appelÃĐ homÃĐostasie
(homÃĐostasie dynamique).
La rÃĐgulation d'un
systÃĻme fait aussi appel à une autre notion, celle de facteur limitant
:
âĒ
Un facteur limitant, aussi appelÃĐ limite de tolÃĐrance,
dÃĐsigne une ressource, un paramÃĻtre ou une condition d'un systÃĻme qui
limite l'ÃĐvolution de tel ou tel processus à l'intÃĐrieur du systÃĻme.
Cela peut Être l'ÃĐlÃĐment sur lequel agit la rÃĐtroaction nÃĐgative,
mais la notion acquiert toute sa pertience lorsqu'elle s'applique à une
ressource rare requise par le systÃĻme considÃĐrÃĐ.
Les systÃĻmes sont exposÃĐs
à de multiples perturbations extÃĐrieures. Deux paramÃĻtres peuvent servir
à mesurer l'ÃĐvolution d'un systÃĻme face aux perturbations : la rÃĐsistance
et la rÃĐsilience.
âĒ
La rÃĐsistance est la capacitÃĐ d'un systÃĻme à maintenir son ÃĐquilibre
malgrÃĐ les perturbations.
âĒ La rÃĐsilience
est fonction de la vitesse à laquelle un systÃĻme retrouve son ÃĐquilibre
aprÃĻs avoir ÃĐtÃĐ perturbÃĐ. Une perturbation peut modifier un systÃĻme
au point qu'il perde entiÃĻrement toute rÃĐsilience. Ce processus peut
conduire à la destruction complÃĻte ou à l'altÃĐration irrÃĐversible
du systÃĻme.
Les systÃĻmes dynamiques
sont d'autant plus aptes à maintenir leur ÃĐquilibre qu'ils sont complexes,
car cela multiplie d'autant les boucles de rÃĐtroaction qui gouvernent
cet ÃĐquilibre.
Les composantes
des ÃĐcosystÃĻmes.
Un ÃĐcosystÃĻme
est un systÃĻme dynamique ouvert, dÃĐfini dans un espace donnÃĐ, qui possÃĻde
une composante vivante (biotique) et une composante non-vivante (abiotique).
L'ÃĐcosphÃĻre est l'ÃĐcosystÃĻme qui renferme tous les autres ÃĐcosystÃĻmes,
à commencer par les diffÃĐrents biomes, qui peuvent eux-mÊmes se dÃĐfinir
comme composÃĐs d'ÃĐcosystÃĻmes de tailles trÃĻs diverses.
On
rend parfois synonymes les mots ÃĐcosystÃĻme et environnement,
mais on doit noter qu'un environnement (qui n'a pas nÃĐcessairement de
composante biotique) doit environner ÂŦ quelque chose Âŧ : on devrait ainsi
parler de l'environnement d'une communautÃĐ biologique, d'un organisme
vivant, etc. C'est l'environnement plus le ÂŦ quelque chose Âŧ qu'il
environne qui pourra donc ÃĐventuellement Être assimilÃĐ Ã un ÃĐcosystÃĻme;
quant à l'ÃĐcosystÃĻme, qui est un systÃĻme ouvert, il a, lui, à coup
sÃŧr un environnement.
Les perturbations extÃĐrieures
auxquelles un ÃĐcosystÃĻme peut Être exposÃĐ sont, par exemple, les effets
du changement climatique ou simplement des alÃĐas mÃĐtÃĐorologiques (variations
des tempÃĐratures et des prÃĐcipitations, sÃĐcheresses, incendies, inondations,
etc.), les effets de l'activitÃĐ humaine (pollution, dÃĐforestation et
autres formes d'anthropisation des sols, introduction d'espÃĻces invasives,
etc.).
On distingue dans
un ÃĐcosystÃĻme :
âĒ
La
biocÃĐnose ou communautÃĐ (ce dernier terme tendant aujourd'hui Ã
supplanter le prÃĐcÃĐdent), renferme la totalitÃĐ de organismes vivants
de l'ÃĐcosystÃĻme. Ces organismes interagissent entre eux et induisent
la structure de l'ÃĐcosystÃĻme.
+ Le
biote. - La biocÃĐnose (en grec, koinÃģs = en commun)
ou communautÃĐ se distingue du biote en ce sens que la biocÃĐnose
dÃĐsigne des ÃĐlÃĐments vivants en interaction, et partant composants d'un
systÃĻme, alors que le mot biote n'a pas ce caractÃĻre dynamique : un biote
est seulement une collection d'Êtres vivants dans un espace dÃĐterminÃĐ.
Pour dÃĐfinir un biote, il suffit d'en ÃĐnumÃĐrer les espÃĻces prÃĐsentes;
pour dÃĐfinir une biocÃĐnose, il faut de surcroÃŪt prÃĐciser qu'elle en
est l'organisation, la diversitÃĐ, la richesse spÃĐcifique.
+ La biomasse.
- Le mot biomasse dÃĐsigne en premier lieu la masse (ou la masse par unitÃĐ
de surface) des organismes vivants prÃĐsents vivants dans une zone ou un
ÃĐcosystÃĻme donnÃĐ, à un moment donnÃĐ. Le mot peut s'appliquer à la
masse de la communautÃĐ toute entiÃĻre ou seulement à celle de certaines
des espÃĻces. D'autres usages du terme se rencontrent : il peut aussi
se rÃĐfÃĐrer seulement à la masse de matiÃĻre organique renfermÃĐe par
les organismes concernÃĐs (on ignore alors la masse de l'eau que referment
ces organismes), Ã la masse de carbone, etc. Dans ces derniers cas,
le terme de biomasse fait ordinairement rÃĐfÃĐrence à la quantitÃĐ de
matiÃĻre organique susceptible d'Être utilisÃĐe comme source d'ÃĐnergie
(bioÃĐnergie).
On dÃĐfinit à l'intÃĐrieur
d'une communautÃĐ biologique diverses composantes-:
+ Selon
les rÃĻgnes qui sont considÃĐrÃĐs, on distinguera : la
phytocÃĐnose
(la flore), la zoocÃĐnose (la faune), la mycocÃĐnose
(les champignons), la microcÃĐnose (les microorganismes), etc.
+ En terme d'espÃĻces
: les organismes d'une mÊme espÃĻce à l'intÃĐrieur d'une communautÃĐ
forment une population.
âĒ Le biotope
(du grec topos = lieu) est la composante de l'ÃĐcosystÃĻme
caractÃĐrisÃĐe par des facteurs abiotiques. Un biotope est ainsi une une
zone limitÃĐe en taille oÃđ vivent des organismes de mÊme espÃĻce ou d'espÃĻces
diffÃĐrentes et oÃđ rÃĻgnent des conditions physiques et chimiques dÃĐfinies
(relativement uniformes dans l'espace et stables dans le temps), telles
que la tempÃĐrature, l'humiditÃĐ, le vent, la lumiÃĻre, le pH, etc. Un
biotope peut Être une souche d'arbre, la forÊt dans laquelle se trouve
cette souche, un ÃĐtang, un verger, une tourbiÃĻre, une vallÃĐe, etc.
+ L'habitat.
- En gÃĐnÃĐral, le terme d'habitat (du latin habitare = vivre)
a un sens similaire à celui de biotope, mais alors que le biotope dÃĐfinit
le lieu de vie d'une communautÃĐ biologique, l'habitat se rÃĐfÃĻre à une
espÃĻce particuliÃĻre, et correspond au lieu de vie d'une population. Ainsi
entendu, l'habitat comprend l'espace que les individus d'une espÃĻce parcourent
pour combler leurs besoins (se nourrir, se cacher, se reposer, se reproduire,
etc.) et peut aussi comprendre leur territoire de migration. Cependant,
certains auteurs anglo-saxons identifient habitat et biotope, quand d'autres
dÃĐfinissent un habitat comme une zone dÃĐterminÃĐe d'un biotope.
âĒ
Niche
ÃĐcologique. - Toutes les espÃĻces ont une niche ÃĐcologique dans l'ÃĐcosystÃĻme.
La notion de niche ÃĐcologique sert à caractÃĐriser la maniÃĻre dont ces
espÃĻces acquiÃĻrent les ressources qui leurs sont nÃĐcessaires et comment
elles interagissent avec les autres espÃĻces de la communautÃĐ.
Il existe divers maniÃĻres d'aborder une niche ÃĐcologique. Par exemple,
a) selon l'habitat dans lequel vit une espÃĻce et les adaptations comportementales
qui l'accompagnent (niche grinnellienne), b) selon la maniÃĻre dont une
espÃĻce ne se contente pas de se dÃĐvelopper dans un environnement, mais
peut aussi modifier l'environnement et son comportement au cours de sa
croissance (niche eltonienne), ou encore c) en fonction de divers paramÃĻtres
statistiques et mathÃĐmatiques qui dÃĐfinissent la maniÃĻre dont une espÃĻce
coexiste avec les autres composantes de la communautÃĐ (niche hutchisonienne).
Les relations trophiques.
La
chaÃŪne trophique.
La circulation de
l'ÃĐnergie et de la matiÃĻre au sein d'une biocÃĐnose est commandÃĐe par
les relations alimentaires qu'y entretiennent ses diffÃĐrentes composantes
(plantes, herbivores, carnivores,
dÃĐcomposeurs), et se dÃĐcrit au travers de la notion de chaÃŪne trophique
( = chaÃŪne alimentaire), qui est
une reprÃĐsentation simplifiÃĐe des interactions entre les diffÃĐrents
organismes dans un ÃĐcosystÃĻme. Une telle chaÃŪne illustre la façon dont
les organismes se nourrissent les uns des autres et comment l'ÃĐnergie
est transfÃĐrÃĐe d'un organisme à un autre. Dans une chaÃŪne alimentaire,
on distingue en particulier, les producteurs, les consommateurs et
les dÃĐcomposeurs :
âĒ Les
producteurs (plantes vertes, algues et certaines bactÃĐries photosynthÃĐtiques),
sont des organismes capables de produire leur propre nourriture par photosynthÃĻse.
Ils convertissent l'ÃĐnergie lumineuse en ÃĐnergie chimique, stockÃĐe sous
forme de matiÃĻre organique (sucres, glucides, etc.).
âĒ Les consommateurs
sont des organismes qui se nourrissent d'autres organismes pour obtenir
de l'ÃĐnergie et des nutriments. Ils peuvent Être classÃĐs en plusieurs
catÃĐgories :
+ Les
herbivores sont des consommateurs primaires qui se nourrissent de plantes
et d'autres producteurs.
+ Les carnivores
sont des consommateurs secondaires ou tertiaires qui se nourrissent principalement
de viande et d'autres animaux.
+ Les omnivores
sont des consommateurs qui se nourrissent à la fois de plantes et d'animaux.
âĒ Les
dÃĐcomposeurs sont des organismes qui dÃĐcomposent la matiÃĻre organique
morte et la dÃĐcomposent en ÃĐlÃĐments nutritifs simples. Ils jouent un
rÃīle essentiel dans le recyclage des ÃĐlÃĐments nutritifs dans l'ÃĐcosystÃĻme
en libÃĐrant les nutriments stockÃĐs dans les dÃĐbris et les organismes
morts.
Les chaÃŪnes trophiques
peuvent Être simples ou complexes en fonction de la diversitÃĐ des espÃĻces
dans l'ÃĐcosystÃĻme et des nombreuses interactions qui s'y produisent.
Le
rÃĐseau trophique.
Les chaÃŪnes alimentaires
ne dÃĐcrivent pas de façon complÃĻte les flux d'ÃĐnergie et de matiÃĻre
dans un ÃĐcosystÃĻme. MÊme quand tous les organismes sont regroupÃĐs dans
les niveaux trophiques appropriÃĐs, on constate que certaines espÃĻces
peuvent se nourrir à partir de plusieurs niveaux trophiques. On a dÃĐjÃ
signalÃĐ le cas des dÃĐcomposeurs, on peut aussi penser aux omnivores qui
peuvent se nourir aussi bien de carnivores, que d'herbivores ou de plantes.
De plus, dans un ÃĐcosystÃĻme, il est rare que l'on n'observe qu'une unique
chaÃŪne alimentaire. Il en existe ordinairement plusieurs, liÃĐes entre
elles pour former une structure complexe. Cela amÃĻne à introduire la
notion de rÃĐseau trophique. Graphiquement, on exprime les relations
trophiques d'une communautÃĐ dans un tel rÃĐseau avec des flÃĻches qui
relient les espÃĻces en fonction de qui mange qui.
-
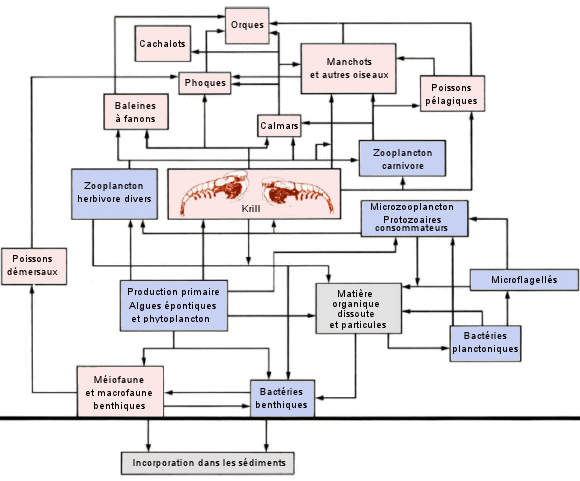
| Le
rÃĐseau trophique dans l'ocÃĐan Antarctique. - Les rÃĐseaux alimentaires
marins polaires ont tendance à Être plus simples que ceux des autres
ÃĐcosystÃĻmes (cela les rend particuliÃĻrement fragiles). Dans l'ocÃĐan
Austral (ainsi d'ailleurs que dans l'ocÃĐan
Arctique), il existe deux types fondamentaux de producteurs primaires
: le phytoplancton pÃĐlagique et les algues qui vivent dans la glace (algues
ÃĐpontiques). Celles-ci sont gÃĐnÃĐralement des espÃĻces benthiques adaptÃĐes
aux faibles intensitÃĐs lumineuses rÃĐgnant sous la surface de la glace.
L'abondant krill (Euphausia superba) constitue le pivot du rÃĐseau trophique
de l'Antarctique car il en est le principal herbivore, et il constitue
aussi une source de nourriture importante pour plusieurs espÃĻces de zooplancton
carnivore, de poissons pÃĐlagiques et de calmars, ainsi que pour les baleines
à fanons, les phoques et les oiseaux de mer qui se nourrissent de plancton.
(D'aprÃĻs
C. Lalli et T. Parsons, Biological oceanography, an introduction,
1993). |
Comme les rÃĐseaux
trophiques peuvent Être trÃĻs compliquÃĐs, on peut les simplifer de diverses
maniÃĻres : par exemple en regroupant les espÃĻces ayant des relations
trophiques similaires dans une communautÃĐ donnÃĐe en grands groupes fonctionnels,
ou encore en isolant une partie du rÃĐseau qui interagit trÃĻs peu
avec le reste de la communautÃĐ.
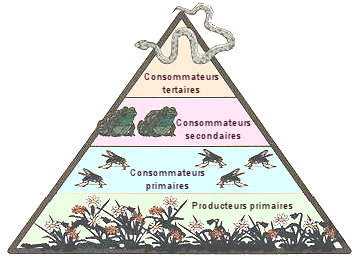
La
pyramide trophique.
On a dit, qu'en
fonction de leur rÃīle de producteur ou de consommateur, les espÃĻces
ou les groupes d'espÃĻces peuvent Être affectÃĐs à diffÃĐrents niveaux
trophiques. Graphiquement, on reprÃĐsente ces niveaux sous forme de bandes
horizontales posÃĐes les uns sur les autres, et dont la longueur est fonction
de grandeurs telles que la biomasse reprÃĐsentÃĐe par tel ou tel niveau,
ou bien la quantitÃĐ d'ÃĐnergie qui peut Être associÃĐe à chacun des
niveaux.
Lors qu'on envisage
l'une ou l'autre de ces caractÃĐristiques, on constate que la biomasse,
le nombre d'organismes ou l'ÃĐnergie sont moindres dans un niveau donnÃĐ
par rapport au prÃĐcÃĐdent, si bien
que le graphique, large à la base, se rÃĐtrÃĐcit à chaque niveau supplÃĐmentaire
pour donner à la figure l'aspect d'une pyramide à degrÃĐs. On appelle
cette figure une pyramide trophique.
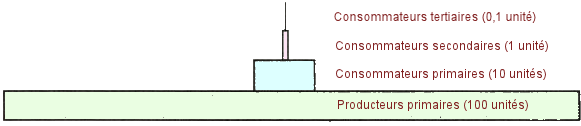
| Pyramide
trophique à quatre niveaux. - Chaque organisme d'une chaÃŪne alimentaire
peut Être placÃĐ dans une pyramide trophique, oÃđ les producteurs sont
situÃĐs en bas et les consommateurs tertiaires en haut. La plupart des
pyramides trophiques ont trois niveaux de consommateurs. Au fur et à mesure
que l'on monte dans la pyramide, on applique la rÃĻgle des 10 %, (voir
ci-dessous) qui stipule qu'Ã chaque niveau 90 % de l'ÃĐnergie utilisable
est perdue dans l'environnement sous forme de chaleur et que seulement
10 % de l'ÃĐnergie utilisable est disponible pour le niveau supÃĐrieur.
Ci-dessous,
une pyramide trophique imagÃĐe et idÃĐalisÃĐe. (Source
: M. Castelluci, Science, 2003). |
La
dÃĐperdition de l'ÃĐnergie.
Les chaÃŪnes alimentaires
d'un rÃĐseau trophique comportent rarement plus de sept maillons (niveaux
trophiques) entre le producteur et n'importe quel prÃĐdateur supÃĐrieur.
La plupart des chaÃŪnes alimentaires sont constituÃĐes quatre maillons,
parfois cinq.
âĒ
La principale raison en est l'inefficacitÃĐ du transfert d'ÃĐnergie
d'un niveau trophique au suivant (hypothÃĻse ÃĐnergÃĐtique). L'ÃĐnergie
est perdue sous forme de chaleur (suite à l'activitÃĐ des organismes)
entre chaque niveau trophique en raison de la deuxiÃĻme loi de la thermodynamique.
Les chiffres peuvent Être diffÃĐrents d'un niveau trophique à un autre
et selon l'ÃĐcosystÃĻme considÃĐrÃĐ, mais on peut estimer qu'en moyenne
seulement 10 % environ de l'ÃĐnergie stockÃĐe dans la matiÃĻre organique
de chaque niveau trophique est convertie en substance organique au niveau
trophique suivant. Il s'ensuit qu'aprÃĻs un nombre limitÃĐ de transferts
trophiques d'ÃĐnergie, la quantitÃĐ d'ÃĐnergie restante dans la chaÃŪne
alimentaire peut ne pas Être assez importante pour soutenir des populations
viables à un niveau niveau trophique supÃĐrieur.
-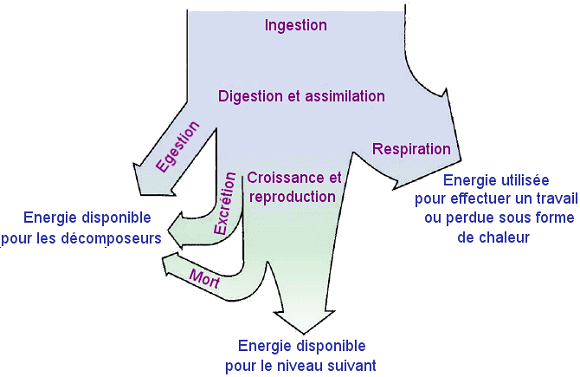 Distribution
de l'ÃĐnergie dans chaque niveau trophique.
Distribution
de l'ÃĐnergie dans chaque niveau trophique.
âĒ Une autre
raison peut aussi Être invoquÃĐe : les chaÃŪnes alimentaires longues sont
moins stables que les chaÃŪnes courtes (hypothÃĻse de stabilitÃĐ dynamique).
Les fluctuations de population aux niveaux trophiques infÃĐrieurs sont
amplifiÃĐes aux niveaux supÃĐrieurs et peuvent conduire à l'extinction
locale des prÃĐdateurs supÃĐrieurs. Pour les grands prÃĐdateurs, plus la
chaÃŪne alimentaire est longue, et plus la rÃĐcupÃĐration des alÃĐas environnementaux
est lente et difficile.
L'amplification
biologique.
IndÃĐpendamment
de la dÃĐperdition d'ÃĐnergie, d'autres phÃĐnomÃĻnes peuvent s'observer
lors du passage d'un niveau trophique au niveau supÃĐrieur. L'un des plus
critiques est l'amplification biologique ou bioamplification,
qui correspond à la concentration croissante de substances persistantes
et toxiques dans les tissus des organismes (bioaccumulation) Ã
la suite de l'ingestion d'espÃĻces du niveau trophique prÃĐcÃĐdent, depuis
les producteurs primaires jusqu'aux consommateurs supÃĐrieurs. De
nombreuses substances sont susceptible d'une telle accumulation pouvant
atteindre le seuil de toxicitÃĐ pour les consommateurs des niveaux trophiques
les plus ÃĐlevÃĐs. Parmi les substances susceptibles d'une bioamplification
problÃĐmatique, on a identifiÃĐ de longue date plusieurs pesticides, Ã
commencer par le DDT(dichlorodiphÃĐnyltrichloroÃĐthane) et le chlordÃĐcone,
ou les BPC (biphÃĐnyles polychlorÃĐs) et les mÃĐtaux lourds (plomb,
mercure, cadmium, etc.).
Les cycles biogÃĐochimiques.
Les processus trophiques
impliquent un flux d'ÃĐnergie et de matiÃĻre. L'ÃĐnergie finit par Être
dissipÃĐe sous forme de chaleur et est perdue à jamais pour les processus
mÃĐtaboliques. Les ÃĐlÃĐments chimiques absorbÃĐs par les organismes vivants
(eau, carbone, azote, phosphore, soufre, etc.) sont appelÃĐs, eux, à Être
recyclÃĐs : ils seront restituÃĐs à la composante abiotique de l'ÃĐcosystÃĻme,
soit du vivant mÊme de ces organismes, soit aprÃĻs leur mort sous l'action
des dÃĐcomposeurs. Il seront alors de nouveau disponibles pour Être utilisÃĐs
par les organismes vivants. Il suivent donc un cycle, appelÃĐ cycle
biogÃĐochimique.
Voici un rapide tour d'horizon des principaux
cycles biogÃĐochimiques. On pourrait encore mentionner les cycles de l'hydrogÃĻne,
du potassium, etc.
Le
cycle de l'eau.
La
disponibilitÃĐ de l'eau influence le rythme de tous les processus à l'oeuvre
dans les ÃĐcosystÃĻmes terrestres, en contrÃīlant la production primaire
et la dÃĐcomposition. De nombreux organismes vivants, comme les plantes,
les animaux et les champignons, dÃĐpendent de la trÃĻs petite quantitÃĐ
d'eau douce de surface, dont le manque peut avoir des effets considÃĐrables.
Les organismes doivent Être capables de s'adapter aux fluctuations de
l'approvisionnement en eau.
L'eau liquide est
la phase physique principale de l'utilisation de l'eau, bien que certains
organismes puissent utiliser la vapeur d'eau. Le gel des eaux souterraines
limite la disponibilitÃĐ de l'eau pour les plantes terrestres. Si l'on
examine les rÃĐserves d'eau sur Terre, on constate que 97,5 % sont des
eaux salÃĐes non potables. Parmi l'eau restante, 99 % est enfermÃĐe dans
le sous-sol sous forme d'eau ou de glace. La quantitÃĐ d'eau dans l'atmosphÃĻre
est minime. Au final, moins de 1% de l'eau douce est facilement accessible
à partir des lacs et des riviÃĻres.
Divers processus
rÃĐgissent le cycle de l'eau. Les principaux sont l'ÃĐvaporation de l'eau
liquide par l'ÃĐnergie solaire qui rÃĐchauffe les ocÃĐans et les autres
eaux de surface, la condensation de la vapeur d'eau dans les nuages et
les prÃĐcipitations, puis l'ÃĐcoulement des eaux de surface et souterraines
qui renvoie l'eau vers les ocÃĐans, complÃĐtant ainsi le cycle :
âĒ
Evaporation / sublimation : Le cycle de l'eau est alimentÃĐ par l'ÃĐnergie
en provenance du Soleil. Cela entraÃŪne l'ÃĐvaporation
(eau liquide vapeur d'eau)
des eaux de surface liquides et la sublimation (glace
vapeur d'eau) de l'eau gelÃĐe, ce qui libÃĻre de grandes quantitÃĐs de
vapeur d'eau dans l'atmosphÃĻre.
âĒ Condensation
/ prÃĐcipitation : au fil du temps, cette vapeur d'eau se condense en nuages
sous forme de gouttelettes liquides ou gelÃĐes et est finalement suivie
de prÃĐcipitations (pluie ou neige),
qui ramÃĻnent l'eau à la surface de la Terre.
âĒ Ecoulement
de l'eau sous la surface : La pluie finit par pÃĐnÃĐtrer dans le sol, oÃđ
elle peut s'ÃĐvaporer à nouveau si elle est proche de la surface, s'ÃĐcouler
sous la surface ou Être stockÃĐe pendant de longues pÃĐriodes.
âĒ Ruissellement
de surface / fonte des neiges : Le ruissellement
de surface, c'est-Ã -dire l'ÃĐcoulement d'eau douce provenant de la pluie
ou de la fonte des glaces, est plus facile à observer.
âĒ Ecoulement
fluvial : Les eaux de ruissellement peuvent ensuite emprunter des cours
d'eau et des lacs pour atteindre les ocÃĐans ou s'ÃĐcouler directement
dans les ocÃĐans eux-mÊmes.
La pluie et le ruissellement
de surface sont les principaux moyens par lesquels les minÃĐraux, notamment
le carbone, l'azote, le phosphore et le soufre, passent de la terre Ã
l'eau.
Le cycle de l'eau
existerait encore en l'absence d'organismes vivants sur la Terre, mais
la transpiration des plantes terrestres qui mobilise un volume d'eau important
lui confÃĻre son caractÃĻre particulier.
Le
cycle du carbone.
Le carbone est le
deuxiÃĻme ÃĐlÃĐment le plus abondant dans les organismes vivants. PrÃĐsent
dans toutes les molÃĐcules organiques, il en forme le squelette et
est aussi au coeur des processus par lesquels l'ÃĐnergie circule à l'intÃĐrieur
des ÃĐcosystÃĻmes. Par ailleurs, liÃĐ Ã l'oxygÃĻne, il forme le CO2
atmosphÃĐrique, qui est le gaz ayant la plus grande part dans l'effet de
serre (L'atmosphÃĻre
de la Terre). Il joue à ce titre un rÃīle pivot dans les ÃĐquilibres
à l'intÃĐrieur de la biosphÃĻre.
La description du
cycle du carbone peut se faire en se plaçant à deux ÃĐchelles temporelles
diffÃĐrentes, celle, courte, des organismes vivants, puis celle de la biosphÃĻre
qui implique de longues durÃĐes-:
âĒ
Les organismes photosynthÃĐtiques (autotrophes) utilisent le dioxyde de
carbone (CO2) atmosphÃĐrique lors de la photosynthÃĻse
et, tout en libÃĐrant de l'oxygÃĻne, convertissent le carbone en formes
organiques qui sont utilisÃĐes par tous les hÃĐtÃĐrotrophes.
La
respiration aÃĐrobie nÃĐcessite de l'oxygÃĻne provenant de l'atmosphÃĻre
ou dissous dans l'eau. Il existe ainsi un ÃĐchange constant d'oxygÃĻne
et de dioxyde de carbone entre les autotrophes (qui ont besoin de carbone)
et les hÃĐtÃĐrotrophes (qui, en plus, ont besoin d'oxygÃĻne).
âĒ Le carbone
utilisÃĐ par les organismes vivants se retrouve ensuite dans le sol et
dans les sÃĐdiments terrestres (oÃđ il forme notamment les grandes rÃĐserves
de combustibles fossiles, qui sont les restes dÃĐcomposÃĐs en anaÃĐrobie
de plantes et qui mettent des millions d'annÃĐes à se former). Sur terre,
le carbone est stockÃĐ dans le sol à la suite de la dÃĐcomposition d'organismes
vivants par des dÃĐcomposeurs ou de l'altÃĐration des roches et des minÃĐraux
terrestres. Ce carbone peut Être aussi lessivÃĐ dans les rÃĐservoirs d'eau
par le ruissellement de surface.
+ L'ÃĐchange
de carbone entre l'atmosphÃĻre et les rÃĐservoirs d'eau influence la quantitÃĐ
de carbone trouvÃĐe en chaque lieu, et chacun affecte l'autre rÃĐciproquement.
Le dioxyde de carbone de l'atmosphÃĻre se dissout dans l'eau et se combine
avec les molÃĐcules d'eau pour former de l'acide carbonique, puis il s'ionise
en ions carbonate et bicarbonate.
La partie du carbone
fixÃĐe dans les roches sÃĐdimentaires
carbonÃĐes telles que le calcaire du fond de l'ocÃĐan est ensuite entraÃŪnÃĐe
dans les profondeurs de la Terre par la subduction des plaques tectoniques.
Le volcanisme sera, plus tard, responsable de son retour dans l'atmosphÃĻre,
bouclant ainsi le cycle.
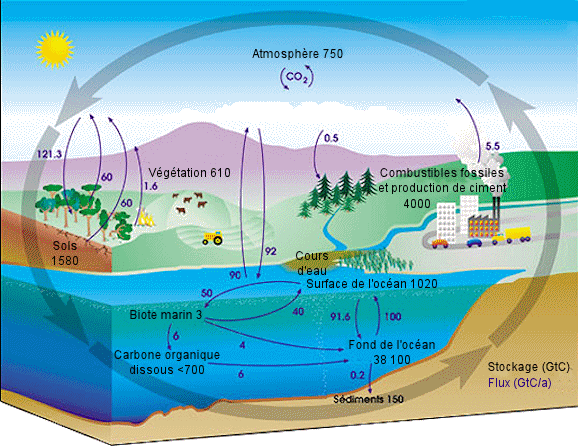
| Le
cycle du carbone. - Chaque annÃĐe, des dizaines de milliards de tonnes
de carbone circulent entre l'atmosphÃĻre, l'hydrosphÃĻre et la lithosphÃĻre.
Les activitÃĐs humaines ajoutent environ 5,5 milliards de tonnes par an
de dioxyde de carbone dans l'atmosphÃĻre. On a mentionnÃĐ en noir les quantitÃĐs
totales de carbone stockÃĐ en gigatonnes de carbone (GtC), et, en violet,
les flux annuels de carbone en gigatonnes de carbone par an (GtC/a).
Source : Nasa Earth Observatory. |
Le
cycle de l'oxygÃĻne.
L'oxygÃĻne est surtout
contenu dans les minÃĐraux de silicate et d'oxydes
de la lithosphÃĻre et du manteau terrestre
(99,5 %). Sa prÃĐsence, comparativement bien moindre, dans l'atmosphÃĻre
terrestre est en lien direct avec les cycles de l'eau et du carbone.
Un petite partie
de cet oxygÃĻne atmosphÃĐrique est issue de la dissociation par le rayonnement
ultraviolet du Soleil des molÃĐcules d'eau et de CO2,
mais l'essentiel provient de la photosynthÃĻse,
qui est une photolyse de l'eau effectuÃĐe par les plantes et le phytoplancton,
et aussi dans une trÃĻs large mesure par une cyanobactÃĐrie marine (le
Prochlorococcus, qui est le plus petit et le plus abondant organisme photosynthÃĐtique
de la planÃĻte), Ã l'origine de la moitiÃĐ de l'oxygÃĻne prÃĐsent dant
l'atmosphÃĻre actuelle de la Terre.
L'oxygÃĻne est fortement
rÃĐactif, il se lie rapidement à d'autres ÃĐlÃĐments (oxydation). Il est
ainsi retirÃĐ en permanence à l'atmosphÃĻre par des rÃĐactions abiotiques
(fixation de l'oxygÃĻne libre par diverses rÃĐactions aux roches de surface)
et biotiques, comme la respiration et la dÃĐcomposition qui aboutissent
à la libÃĐration dans l'atmosphÃĻre de dioxyde de carbone. Si l'atmosphÃĻre
parvient à maintenir un niveau de 21% d'oxygÃĻne, elle le doit seulement
à l'activitÃĐ photosynthÃĐtique des organismes vivants qui y injectent
aussi en permanence ce gaz.
Le
cycle de l'azote.
L'azote est un composant
des acides aminÃĐs, des protÃĐines et des acides
nuclÃĐiques, et est un ÃĐlÃĐment nutritif essentiel et particuliÃĻrement
important pour l'ÃĐtude de la dynamique des ÃĐcosystÃĻmes, puisque de nombreux
processus, tels que la production primaire et la dÃĐcomposition, sont limitÃĐs
par l'apport d'azote disponible.
Le principal rÃĐservoir
d'azote est l'atmosphÃĻre, composÃĐe à 78 % d'azote gazeux (N2).
Les autres rÃĐservoirs sont dans le sol, la biomasse, les sÃĐdiments au
fond des lacs, des cours d'eau et des ocÃĐans, ou encore, sous forme dissoute,
les eaux de surface et souterraines.
Les producteurs primaires,
tels que les plantes et le phytoplancton, qui ordinairement font entrer
les nutriments dans la chaÃŪne trophique ne sont pas capables d'incorporer
l'azote de l'atmosphÃĻre. La principale voie d'entrÃĐe de l'azote dans
les ÃĐcosystÃĻmes est donc plutÃīt la fixation du N2
par des bactÃĐries (bactÃĐries nitrifiantes),
telles, par exemple, les Rhizobium, qui vivent en symbiose dans les nodules
racinaires des lÃĐgumineuses, ou les Azobacter,
qui sont des bactÃĐries libres. Ces bactÃĐries lient l'azote à l'oxygÃĻne
pour former l'ion nitrate (NO3-), qui associÃĐ
à divers anions (ions positifs), forme les nitrates proprement dits. Ces
composÃĐs azotÃĐs organique peuvent alors Être absorbÃĐs aisÃĐment par
les plantes.
Dans la suite du
cycle, l'azote sera rÃĐintroduit dans l'atmosphÃĻre par d'autres bactÃĐries
ou par des champignons sous forme d'azote gazeux. Ce processus se dÃĐroule
en trois ÃĐtapes dans les systÃĻmes terrestres : ammonification, nitrification
et dÃĐnitrification.
âĒ
L'ammonification convertit les dÃĐchets azotÃĐs d'animaux vivants ou de
restes d'animaux morts en ammonium (NH4+)
par certaines bactÃĐries et champignons.
âĒ La nitrification
est la conversion de NH4+ en NO3-.
L'ammonium est d'abord converti en nitrites (NO2-)
par des bactÃĐries nitrifiantes, telles que Nitrosomonas. Les nitrites
sont ensuite convertis en nitrates (NO3-)
par des organismes similaires.
âĒ La dÃĐnitrification
intervient dans des conditions anaÃĐrobies-:
les bactÃĐries nitrifiantes, telles que Pseudomonas et Clostridium, utilisent
du NO3- au lieu de l'O2
pour leur mÃĐtabolisme et libÃĻrent du N2 (azote gazeux)
qui peut rÃĐintÃĐgrer l'atmosphÃĻre.
Le cycle de l'azote
ocÃĐanique se dÃĐroule de façon trÃĻs similaire par l'intermÃĐdiare aussi
de bactÃĐries. Une partie de cet azote tombe au fond de l'ocÃĐan sous forme
de sÃĐdiments que les mouvements gÃĐologiques finissent par faire revenir
en surface, oÃđ l'azote prÃĐsent peut ÃĐventuellement Être rÃĐinjectÃĐ
dans les ÃĐcosystÃĻmes terrestres.
Cycle
du phosphore.
Le phosphore est
un constituant essentiel des acides nuclÃĐiques,
des phospholipides, de l'ATP et d'autres molÃĐcules
de stockage d'ÃĐnergie; sous la forme de phosphate de calcium, il constitue
les composants de soutien des os. NÃĐcessaire à la
croissance des Poissons, le phosphore est souvent le nutriment limitant
dans les ÃĐcosystÃĻmes aquatiques.
Aucun gaz atmosphÃĐrique
ne contient du phosphore (cet ÃĐlÃĐment n'est ÃĐventuellement prÃĐsent
dans l'atmosphÃĻre que sous forme de poussiÃĻre minÃĐrale, d'aÃĐrosols
ou de cendres volcaniques). En renvanche, le phosphore se rencontre en
grandes quantitÃĐs dans le sol ou dissous dans les ocÃĐans, ainsi
que dans les organismes vivants. Mais ce sont les roches sÃĐdimentaires
d'origine marine qui constituent le principal rÃĐservoir de phosphore :
il y est issu des corps des organismes ocÃĐaniques et de leurs excrÃĐtions.
La seule forme de
phosphore matiÃĻre inorganique d'importance biologique est l'ion phosphate
(PO4+), que les plantes absorbent et utilisent
pour synthÃĐtiser des composÃĐs organique ensuite distribuÃĐs dans tout
le rÃĐseau trophique. Le phosphate retourne au sol ou à l'eau par la dÃĐcomposition
de
la biomasse ou l'excrÃĐtion par les consommateurs. L'humus et les particules
de sol lient alors les phosphates, les rendant de nouveau disponibles pour
les producteurs primaires.
Le cycle du phosphore
a ainsi tendance à Être largement localisÃĐ dans les ÃĐcosystÃĻmes. Mais
le parcours du phosphore peut aussi Être beaucoup plus long. Une partie
des phosphates contenus dans les roches est lessivÃĐe par les eaux souterraines
et de surface et atteint finalement les ocÃĐans. Le retour du phosphate
de l'ocÃĐan vers la terre et à travers le sol est ensuite extrÊmement
lent : l'ion phosphate a un temps de sÃĐjour ocÃĐanique
moyen compris entre 20.000
et 100.000 ans.
Cycle
du soufre.
En tant que composant
de la cystÃĐine (un acide aminÃĐ), le soufre est impliquÃĐ, au sein des
protÃĐines,
dans la formation de liaisons disulfure, qui contribuent à dÃĐterminer
leur repliement tridimensionnel, et donc leurs fonctions. Par ailleurs,
certains ÃĐcosystÃĻmes reposent sur des chimioautotrophes utilisant le
soufre (sous forme de sulfates) comme source d'ÃĐnergie biologique.
PrÃĐsent dans l'atmosphÃĻre
sous forme de dioxyde de soufre (SO2), le soufre y
pÃĐnÃĻtre de trois maniÃĻres : par la dÃĐcomposition de molÃĐcules organiques,
par l'activitÃĐ volcanique et les ÃĐvents gÃĐothermiques, et par la combustion
de combustibles fossiles par les humains. Il se dÃĐpose ensuite sur la
terre aussi de trois maniÃĻres principales :
âĒ
Lorsque la pluie tombe, le dioxyde de soufre atmosphÃĐrique se trouve dissous
sous forme d'acide sulfureux (H2SO3).
âĒ Le soufre
peut ÃĐgalement tomber directement de l'atmosphÃĻre.
âĒ De plus,
l'altÃĐration des roches contenant du soufre libÃĻre du soufre dans le
sol. Ces roches proviennent de sÃĐdiments ocÃĐaniques qui sont dÃĐplacÃĐs
vers la terre par le soulÃĻvement gÃĐologique de ces sÃĐdiments.
Les ÃĐcosystÃĻmes terrestres
utilisent ces sulfates du sol sous forme liquide. L'ion sulfate (SO4--)
est rÃĐduit par les plantes et les autres producteurs primaires qui l'incorporent
à leurs protÃĐines sous forme solide. Ensuite, les consommateurs des ces
plantes l'intÃĻgrent à leurs protÃĐines. Lors de la mort et de la
dÃĐcomposition de ces organismes, le soufre rÃĐduit des protÃĐines est
oxydÃĐ par les bactÃĐries et de nouveau assimilable par les plantes, ou
bien il est rejetÃĐ dans l'atmosphÃĻre sous forme de sulfure d'hydrogÃĻne
(H2S).
Le soufre pÃĐnÃĻtre
dans l'ocÃĐan par le ruissellement de la terre, par les retombÃĐes atmosphÃĐriques
et par les ÃĐvents gÃĐothermiques sous-marins.
Les
effets de l'activitÃĐ humaine sur les cycles biogÃĐochimiques.
En raison des capacitÃĐs
technologiques des humains et des systÃĻmes de reprÃĐsentation dans lesquels
ils inscrivent leurs existences individuelles, qui induisent chez eux des
besoins qui vont bien au-delà de la dimension simplement biologique, les
humains ont, sur l'ÃĐcosphÃĻre, un impact collectif bien plus grand et
bien plus variÃĐ que celui des autres animaux. On se contentera ici d'ÃĐvoquer
l'impact des activitÃĐs humaines sur les cycle biogÃĐochimiques.
Afin d'accroÃŪtre
la disponibilitÃĐ de l'eau à leur bÃĐnÃĐfice, les humains modifient
le cycle de l'eau de diverses maniÃĻres (collecte d'eaux souterraines,
barrages sur les cours d'eau, irrigation, recours au dessalement pour obtenir
de l'eau potable à partir de l'ocÃĐan, etc).
-

| Le
fond assÃĐchÃĐ de la mer d'Aral en OuzbÃĐkistan. - Autrefois alimentÃĐ
en eau par deux fleuves importants, l'Amou-Daria et le Syr-Daria, ce lac
d'Asie centrale est aujourd'hui en grande partie assÃĐchÃĐ. Cette catastrophe
ÃĐcologique majeure est due au du dÃĐtournement des deux fleuves, dans
la seconde moitiÃĐ du XXe siÃĻcle, pour servir à l'irrigation de grandes
plantations de coton. Photo : Anton Ruiter; licence
; Creative Commons. |
Les humains interviennent
de maniÃĻre plus cruciale encore dans le cycle du carbone, en partie du
fait de l'ÃĐlevage de bovins, source mÃĐthane (CH4),
mais principalement depuis le dÃĐbut de l'ÃĻre industrielle, par l'injection
dans l'atmosphÃĻre de quantitÃĐs massives de dioxyde de carbone issu de
la combustion des combustibles fossiles. La prÃĐsence croissante dans l'atmosphÃĻre
de CH4 et CO2, qui sont des gaz
à effet de serre, affecte profondÃĐment le climat et bouleverse tous les
ÃĐcosystÃĻmes. Les ocÃĐans, qui absorbent environ un quart du CO2
atmosphÃĐrique, contribuent à limiter jusqu'à un certain point cet effet
de serre, mais en contrepartie ils s'acidifient, ce qui met en pÃĐril les
ÃĐcosystÃĻmes marins. Lacidification des ocÃĐans a augmentÃĐ de 26.%
depuis le dÃĐbut de l'ÃĻre industrielle et ne fait que s'accÃĐlÃĐrer.
La combustion de
combustibles fossiles libÃĻre aussi des oxydes d'azote (oxyde nitreux,
N2O, en particulier) qui sont aussi des gaz à effet
de serre. Mais les rejets de composÃĐs azotÃĐs par les activitÃĐs humaines
sont surtout reliÃĐs à l'agriculture. Pour faciliter la croissance et
la productivitÃĐ des plantes cultivÃĐes, on leur apporte souvent des nutriments
en abondance, sous forme d'engrais. Ces engrais contiennent notamment de
l'azote (nitrates), du phosphore (phosphates) et du potassium, et les cycles
de ces ÃĐlÃĐments sont eux aussi fortement affectÃĐs. Les excÃĻs de ces
ÃĐlÃĐments pÃĐnÃĻtrent dans les sols ou sont vÃĐhiculÃĐs par le ruisselement
de surface jusqu'aux riviÃĻres, aux lacs et aux ocÃĐans. Ils provoquent
alors une croissance excessive de micro-organismes (processus d'eutrophisation),
qui ÃĐpuisent l'oxygÃĻne dissous, ce qui entraÃŪne la mort de la faune
de nombreux ÃĐcosystÃĻmes. Ce processus est responsable des zones mortes
(zones incapables de supporter des organismes vivants) dans les lacs et
à l'embouchure de nombreux grands fleuves. On estime que ces zones mortes,
dont certaines existent aussi en plein ocÃĐan (peut-Être, dans ce cas,
pour des raisons liÃĐes au rÃĐchauffement climatique) reprÃĐsentent aujourd'hui
dans le monde une superficie de l'ordre de 250 000 kmÂē, soit plus que
celle du Royaume-Uni et prÃĻs de la moitiÃĐ
de la superficie de la France mÃĐtropolitaine
Enfin, le cycle du
soufre est lui aussi fortement impactÃĐ par les activitÃĐs humaines, Ã
commencer à par le recours aux combustibles fossiles (pÃĐtrole, gaz naturel,
charbon) qui libÃĻre dans l'atmosphÃĻre du dioxyde de soufre (SO2)
et de sulfure d'hydrogÃĻne (H2S). L'eau de pluie
tombant sur le sol à travers le sulfure d'hydrogÃĻne et le dioxyde de
soufre les transforme en acide sulfureux (H2SO3).
Il en rÃĐsulte des pluies acides toxiques qui abaissent le pH des lacs
(ce qui tue une grande partie de la faune rÃĐsidente); les pluies acides
impactent ÃĐgalement directement les humains en affectant leur santÃĐ et
en dÃĐgradant chimiquement les bÃĒtiments. (Ajoutons que les plus acides
peuvent avoir d'autres origines, notamment du fait de rejets de composÃĐs
azotÃĐs, tels que l'acide nitrique, HNO3).
Les communautÃĐs biologiques
On nomme communautÃĐ
(= biocÃĐnose) l'ensemble des espÃĻces qui cohabitent et interagissent
au sein d'un mÊme ÃĐcosystÃĻme. Les communautÃĐs sont des entitÃĐs complexes
dÃĐfinies par leur structure (les types et le nombre d'espÃĻces) et leur
dynamique (comment elles changent au fil du temps).
Les interactions
biologiques.
Dans une communautÃĐ, les interactions
entre individus de deux espÃĻces (individus hÃĐtÃĐrospÃĐcifiques) ou d'une
mÊme espÃĻce (individus conspÃĐcifiques) peuvent prendre plusieurs formes.
On parle de neutralisme dans le cas oÃđ aucune des deux ne subit
ni de prÃĐjudice ni de bÃĐnÃĐfice du fait de l'interaction. L'amensalisme
correspond à la situation dans laquelle la relation cause un prÃĐjudice
à l'une, mais a un effet neutre pour l'autre. On parle enfin d'antagonisme
quand l'interaction bÃĐnÃĐficie à une espÃĻce et est prÃĐjudiciable Ã
l'autre. On parle de facilitation quand au moins l'un
des espÃĻces tire bÃĐnÃĐfice de l'interaction sans porter prÃĐjudice Ã
l'autre (mutualisme, commensalisme).
Une interaction entre
deux organismes est dite obligatoire lorsque au moins l'un des deux
organismes ne pourrrait vivre l'un sans interaction avec l'autre. Elle
est dit facultative, quand l'un ces organismes sont capables
de survivre sans cette interaction. Certaines interactions sont de courte
durÃĐe (par exemple la prÃĐdation, qui est instantanÃĐe, la pollinisation;
la zoocorie), d'autres de longue durÃĐe, comme le mutualisme, le parasitisme,
la compÃĐtition, le neutralisme, etc.
Les principaux
types d'interactions biologiques
| EspÃĻce |
Antagonisme |
Mutualisme |
Amensalisme |
Commensalisme |
Neutralisme |
| I |
+ |
+ |
= |
= |
= |
| II |
- |
+ |
- |
+ |
= |
| Le
signe + signifie que l'interaction est bÃĐnÃĐfique pour les individus de
l'espÃĻce concernÃĐe; le signe - signifie que l'interaction a des effets
nÃĐfastes sur ces individus; le signe = signifie que l'impact de l'interaction
n'est ni positif ni nÃĐgatif. |
Antagonisme
(+ -).
Les interactions
antagonistes sont celles dans lesquelle l'interaction favorise un organisme
ou une espÃĻce au dÃĐtriment d'un autre organisme ou d'une autre espÃĻce.
Le comportement associÃĐ Ã ce type d'interaction est appelÃĐ exploitation
(d'une espÃĻce par l'autre). Les espÃĻces prÃĐjudiciÃĐes ont souvent dÃĐveloppÃĐ
au cours de leur ÃĐvolution diverses stratÃĐgies de dÃĐfense qui peuvent
Être mÃĐcaniques, chimiques, comportementales, etc., tels sont le mimÃĐtisme,
les piquants, la toxicitÃĐ, les carapaces, la fuite ou, au contraire, l'immobilitÃĐ,
etc.
âĒ
La
prÃĐdation est l'interaction entre animaux,
dans laquelle un individu d'une espÃĻce (le prÃĐdateur) se nourrit, aprÃĻs
capture, d'un individu d'une autre espÃĻce (la proie), qui ne survit pas
à l'interaction.
âĒ L'herbivorÃe
(ou phytophagie) est l'interaction dans laquelle un animal ne nourrit
d'un vÃĐgÃĐtal. Elle peut se dÃĐcliner de diverses
façons, selon les vÃĐgÃĐtaux ou les parties de vÃĐgÃĐtal consommÃĐs
: herbivorie proprement dite (herbe); phyllophagie ou folivorie (feuilles);
frugivorie (fruits); granivorie (graines).
Chaque terme a son pendant dans la qualification de l'animal selon son
mode d'alimentation (herbivore, phyllophage; frugivore, granivore, etc.).
âĒ Le parasitisme
est une interaction dans laquelle l'organisme d'une espÃĻce (le parasite)
vit à la surface (ectoparasitisme), à l'intÃĐrieur (endoparasitisme)
ou en partie en surface et en partie à l'intÃĐrieur (mÃĐsoparasitisme)
d'un organisme d'une autre espÃĻce (l'hÃīte) pour en tirer un bÃĐnÃĐfice,
gÃĐnÃĐralement à son dÃĐtriment (parfois pouvant mÊme entraÃŪner la mort
de l'hÃīte). Le bÃĐnÃĐfice tirÃĐ peut Être divers. Le parasite peut se
nourrir grÃĒce à son hÃīte, ou avoir besoin de ce dernier pour se reproduire,
comme dans le cas des virus. Cette interaction
peut Être facultative pour certaines espÃĻces de parasites, mais elle
obligatoire pour d'autres.
Le
coopÃĐration et le mutualisme (+ +).
La coopÃĐration
est une interaction ayant des effets bÃĐnÃĐfiques rÃĐciproques pour des
organismes d'espÃĻces diffÃĐrentes ou appartenant à la mÊme espÃĻce.
Dans le cas de coopÃĐration entre deux espÃĻces distinctes, on parle de
mutualisme.
Le mutualisme est une interaction à long terme. La symbiose et le protocoopÃĐration
sont deux types de mutualisme :
âĒ
La
symbiose est une forme de mutualisme entre deux espÃĻces dans laquelle
tous ou quelques-un des individus concernÃĐs sont bÃĐnÃĐficiaires. C'est
une interaction obligatoire pour l'une des espÃĻce concernÃĐes, Ã au moins
un stade de sa vie. La symbiose est dite unilattÃĐrale quand le
caractÃĻre obligatoire ne concerne que l'une des espÃĻces; elle est bilatÃĐrale
quand l'interaction est obligatoire pour les deux. On appelle holobionte
le tout constituÃĐ par des organismes vivant en symbiose.
âĒ La protocoopÃĐration
est une interaction rendant sont mutuellement bÃĐnÃĐficiaires les individus
ou les populations qu'elle concerne, sans que cette relation soit essentielle
pour l'une ou l'autre. Deux exemples : La pollinisation d'une plante
phanÃĐrogame par un insecte, la mycorhyse entre entre
un champignon et les racines d'une plante.
Amensalisme
(= -).
L'amensalime correspond
à un type d'interaction de longue durÃĐe entre organismes, qui n'a ni
bÃĐnÃĐfice ni prÃĐjudice pour l'un, mais qui est prÃĐjudiciable à l'autre.
On distingue principalement deux formes d'amensalisme, l'allÃĐlopathie
nÃĐgative et la
compÃĐtition :
âĒ
L'allÃĐlopathie
nÃĐgative est une interaction biochimique entre deux organismes, dans
laquelle un organisme est nÃĐfaste à un autre en produisant et diffusant
certains composÃĐs chimiques.
âĒ
La
compÃĐtition est une interaction dans laquelle la prÃĐsence d'un organisme
dans un une zone rÃĐduit ou prive complÃĻtement un autre organisme de ressources
vitales (nourriture, lumiÃĻre), qui sera dÃĻs lors exclu de cet habitat.
De ce point de vue la compÃĐtition, qui peut aussi Être prÃĐjudiciable
aux deux individus ou espÃĻces en prÃĐcence (- -) peut ÃĐgalement Être
rangÃĐe parmi les antagonismes.
Le
principe d'exclusion compÃĐtitive. - Ce principe stipule que, dans un habitat
donnÃĐ, deux espÃĻces ne peuvent pas occuper la mÊme niche ÃĐcologique
: diffÃĐrentes espÃĻces ne peuvent pas coexister dans une communautÃĐ si
elles sont en concurrence pour les mÊmes ressources.

| Un
Renard et un Blaireau occupÃĐs à se nourrir de prunes tombÃĐes de l'arbre.
- Ces deux espÃĻces, classÃĐes parmi les Carnivores, sont omnivores en
pratique. Elles peuvent partager le mÊme habitat, oÃđ elles sont en compÃĐtition
pour l'accÃĻs à la nourriture (occasionnellement de façon trÃĻs flagrante),
mais, malgrÃĐ des chevauchements possibles pour lesquels il existe une
compÃĐtition, elles n'occupent pas exactement la mÊme niche ÃĐcologique
(partitionnement de niche), ce qui assure leur coexistence durable. Photo
: ÂĐ Serge Jodra, 2022. |
Le
commensalisme (= +).
Contrairement Ã
l'amensalisme, le commensalisme (au sens large) dÃĐsigne une des interactions
entre organismes qui, si elles n'ont ni bÃĐnÃĐfice ni prÃĐjudice pour l'un
(appelÃĐ l'hÃīte), ont un effet bÃĐnÃĐfique sur l'autre (le commensal,
l'inquilin, le phoronte, l'ÃĐpibionte, etc).
âĒ
Commensalisme
proprement dit. - Au sens propre, le commensalisme dÃĐsigne
les interactions dans lesquelles une espÃĻce prÃĐlÃĻve sa nourriture Ã
partir d'une autre. L'ÃĐtymologie (le mot vient du latin cum = avec
et mensa = table), rappelle que le commensal est celui qui mange
"à la mÊme table" que son hÃīte.
On peut y rattacher
d'autres interactions telles que :
âĒ
L'inquilinisme,
qui concerne les situations dans lequelles une espÃĻce est hÃĐbergÃĐe par
un hÃīte auquel elle ne nuit ni de bÃĐnÃĐficie. Par exemple, quand un oiseau
utilise le nid d'un autre aprÃĻs que celui-ci l'ait
abandonnÃĐ.
âĒ L'allÃĐlopathie
positive, qui est une interaction biochimique entre deux organismes,
dans laquelle un organisme bÃĐnÃĐficie à un autre en produisant certains
composÃĐs chimiques. LallÃĐlopathie, qu'elle soit positive ou nÃĐgative,
concerne essentiellement les plantes et les micro-organismes.
âĒ L'ÃĐpibiose,
qui correspond à l'interaction entre une espÃĻce et une autre espÃĻce
qui lui sert de support (cas des mousses , des
lichens,
des orchidÃĐes). Contrairement au parasite,
l'ÃĐpibionte ne prÃĐlÃĻve rien au dÃĐtriment de son hÃīte.
âĒ La phorÃĐsie,
qui renvoie à des situations oÃđ une espÃĻce se sert d'une autre comme
moyen de transport. Exemple, les rÃĐmoras qui vivent accrochÃĐs aux requins.
âĒ La zoocorie
est une interaction qui peut Être rapprochÃĐe de la prÃĐcÃĐdente, et par
laquelle un animal transporte les graines des plantes et contribue ainsi
à la dissÃĐmination des semences. Dans l'exozoocorie, les graines
adhÃĻrent à la peu de l'animal; dans l'endozoocorie, les graines
sont absorbÃĐes par l'animal, puis rejetÃĐes dans ses excrÃĐments.
âĒ La thanatochrÃĻse
est l'utilisation de restes, d'excrÃĐments, de squelettes ou de cadavres
d'autres espÃĻces afin de se protÃĐger ou de les utiliser comme outils.
Par exemple, les pagures (bernard-l'hermite), qui utilisent des coquilles
de gastÃĐropodes pour protÃĐger leur abdomen.
La biodiversitÃĐ.
La biodiversitÃĐ
ou diversitÃĐ biologique dÃĐfinit la complexitÃĐ biologique d'une communautÃĐ.
Cette diversitÃĐ a deux composantes :
âĒ
La
richesse en espÃĻces (richesse spÃĐcifique), qui est le nombre d'espÃĻces
diffÃĐrentes vivant dans dans une zone particuliÃĻre, un habitat,
un biome ou mÊme la biosphÃĻre dans son entier. La plus grande richesse
en espÃĻces s'observe dans les ÃĐcosystÃĻmes proches de l'ÃĐquateur
(tempÃĐratures plus chaudes, grandes quantitÃĐs de prÃĐcipitations, faible
saisonnalitÃĐ); Ã l'inverse, la plus faible richesse en espÃĻces se rencontre
aux trÃĻs hautes latitudes (tempÃĐratures trÃĻs
basses, faibles prÃĐcipitations).
âĒ L'abondance
relative des espÃĻces (rÃĐgularitÃĐ spÃĐcifique), dans une zone particuliÃĻre,
qui est le nombre d'individus d'une espÃĻce par rapport au nombre total
d'individus de toutes les espÃĻces dans cette zone. Si l'on a deux communautÃĐs
de mÊme richesse en espÃĻces, on considÃĻre que la biodiversitÃĐ est plus
grande dans celle oÃđ l'abondance relative de chaque espÃĻce est la plus
grande; il y a moins de biodiversitÃĐ lorsque certaines espÃĻces sont trÃĻs
reprÃĐsentÃĐes et d'autres trÃĻs peu.
La richesses et l'abondance
relative des diffÃĐrentes espÃĻces qui composent une communautÃĐ sont importantes
pour l'intÃĐgritÃĐ et souvent mÊme la survie de la communautÃĐ (on a dit
plus haut que plus un systÃĻme est complexe et plus il dispose de leviers
sur lesquels il peut agir pour se rÃĐguler). Les communautÃĐs diversifiÃĐes
protÃĻgent les ÃĐcosystÃĻmes en fournissant des services tels que l'ombre,
la dÃĐgradation des dÃĐchets et la production d'oxygÃĻne.
Le succÃĻs et la
survie d'une espÃĻce dÃĐpendent aussi de sa diversitÃĐ gÃĐnÃĐtique, c'est-Ã -dire
de la variÃĐtÃĐ et de la frÃĐquence relative des diffÃĐrents allÃĻles
dans son patrimoine gÃĐnÃĐtique. La diversitÃĐ gÃĐnÃĐtique peut Être essentielle
pour qu'une espÃĻce puisse s'adapter à des environnements changeants.
EspÃĻces
au rÃīle particulier.
Au sein d'une communautÃĐ,
certaines espÃĻces exercent un fort contrÃīle sur la structure de la communautÃĐ,
en particulier sur la composition des espÃĻces prÃĐsentes, leur abondance
relative et sur la diversitÃĐ en gÃĐnÃĐral. Soit en raison de leur abondance,
soit parce qu'elles jouent un rÃīle fondamental dans la dynamique de cette
communautÃĐ, l'impact de ces espÃĻces peut s'exercer par des interactions
trophiques ou par leurs influences sur le milieu physique. On distingue
ainsi :
âĒ
Les
espÃĻces fondatrices. - Les espÃĻces fondatrices sont considÃĐrÃĐes
comme le socle sur lequel se constitue une communautÃĐ; ce sont celles
qui ont la plus grande influence sur sa structure globale. Elles possÃĻdent
souvent l'abondance relative la plus ÃĐlevÃĐe. Ce sont gÃĐnÃĐralement les
producteurs primaires : des organismes qui apportent la majeure partie
de l'ÃĐnergie à la communautÃĐ. Elles n'exercent leur influence en provoquant
des changements physiques dans l'environnement (soit par leur comportement,
soit par leur importante biomasse) pour produire et maintenir des habitats
qui profitent aux autres organismes qui les utilisent. En modifiant la
structure ou la dynamique de l'environnement, les espÃĻces fondatrices
agissent comme des facilitateurs, avec des effets positifs sur la survie
et la reproduction de certaines autres espÃĻces de la communautÃĐ.
âĒ
L'espÃĻce
dominante. - On nomme espÃĻce dominante d'une communautÃĐ celle qui
y exerce le plus fort contrÃīle sur la prÃĐsence et la distribution d'autres
espÃĻces. C'est ordinairement la plus abondante ou celle qui reprÃĐsente
la plus importante biomasse. L'espÃĻces fondatrice peut ainsi parfois Être
l'espÃĻce dominante. Le rÃīle dominant d'une espÃĻce peut lui venir de
ce qu'elle est plus compÃĐtitive dans l'exploitation de ressources limitÃĐes
telles que l'eau ou les nutriments. Elle peut Être dominante aussi parce
qu'elle est plus efficace pour ÃĐviter la prÃĐdation ou les consÃĐquences
des maladies.
âĒ Les
espÃĻces clÃĐs. - Une espÃĻce clÃĐ est une espÃĻce dont la prÃĐsence
est essentielle au maintien de la biodiversitÃĐ au sein d'un ÃĐcosystÃĻme
et au maintien de la structure d'une communautÃĐ ÃĐcologique. Contrairement
aux espÃĻces dominantes, les espÃĻces clÃĐs n'ont pas besoin d'Être abondantes
dans une communautÃĐ. Elles exercent un fort contrÃīle sur la structure
de la communautÃĐ d'abord parce qu'elles y jouet un rÃīle ou y occupent
une niche ÃĐcologique fondamentale.
Les successions ÃĐcologiques.
Tous les espaces
qui offrent des conditions de vie propices à des organismes vivants sont
occupÃĐs. Des communautÃĐs s'y sont constituÃĐes qui y ont acquis une structure
stable. Mais il peut arriver que la composition et la structure communautaire
soient notablement altÃĐrÃĐes, surtout quand toute la vÃĐgÃĐtation existante
dans la zone est dÃĐmantelÃĐe. Cela peut survenir à la suite de perturbations
environnementales telles que des ÃĐruptions volcaniques, des sÃĐismes,
des tempÊtes, des incendies, la rÃĐgression de glaciers, l'assÃĻchement
de zones humides, etc. Les effets du changement climatique et l'intervention
humaine jouent ici un rÃīle majeur. L'ÃĐquilibre est rompu parfois temporairement,
parfois dÃĐfinitivement. Dans ce cas, la zone vidÃĐe totalement ou partiellement
des espÃĻces qui y vivaient peut Être colonisÃĐe par de nouvelles espÃĻces,
qui d'ailleurs pourront Être progressivement remplacÃĐes par d'autres
qui, Ã leur tour, seront possiblement remplacÃĐes par d'autres encore,
jusqu'à ce que s'ÃĐtablisse à nouveau un ÃĐcosystÃĻme avec une communautÃĐ
à l'ÃĐquilibre. Ce processus d'apparition et de disparition sÃĐquentielles
au fil du temps d'espÃĻces dans une communautÃĐ est appelÃĐ succession
ÃĐcologique. L'ÃĐtat final d'une succession est appelÃĐ son climax.
Les successions peuvent
revÊtir des caractÃĻres diffÃĐrents, selon lien qui existe entre la premiÃĻre
espÃĻces installÃĐe et la deniÃĻre. On peut ici distinguer trois modalitÃĐs
principales :
âĒ
Les espÃĻces prÃĐcoces facilitent l'apparition des espÃĻces plus tardives,
leur rendant l'environnement plus favorable (par exemple, en augmentant
la fertilitÃĐ du sol).
âĒ Les premiÃĻres
espÃĻces inhibent l'implantation des espÃĻces ultÃĐrieures, mais pas suffisamment
pour l'empÊcher.
âĒ Les premiÃĻres
espÃĻces sont totalement indÃĐpendantes des suivantes : elles n'aident
ni n'entravent leur colonisation.
Succession
primaire.
Dans la succession
primaire, les terres nouvellement exposÃĐes ou nouvellement formÃĐes sont
vides de toute espÃĻce vivante et sont colonisÃĐes entiÃĻrement pas de
nouvelles espÃĻces. Les premiers organismes à s'installer sont les procaryotes
autotrophes.Des organismes photosynthÃĐtiques macroscopiques (lichens et
mousses) se dÃĐveloppent ensuite à partir de spores apportÃĐs par le vent.
Le sol se transforme progressivement, son pH ÃĐvolue, Ã mesure que les
roches se dÃĐtÃĐriorent sous l'effet des intempÃĐries, et que la matiÃĻre
organique s'accumule à partir des restes en dÃĐcomposition. Il devient
ainsi propice à des installations nouvelles. Des herbes commencent Ã
pousser et recouvrent les mousses et les lichens. Ces toutes premiÃĻres
espÃĻces, qui contribuent à enrichir de plus en plus le sol, sont connues
sous le nom d'espÃĻces pionniÃĻres. D'autres espÃĻces moins rÃĐsistantes
poussent et remplacent ÃĐventuellement les espÃĻces pionniÃĻres : ce peut
Être des buissons et des arbustes, dont les graines ont ÃĐtÃĐ ÃĐtÃĐ apportÃĐes
par les animaux qui commencent à occuper ce nouvel espace. Certaines de
ce ses nouvelles espÃĻces, apportent peut-Être aussi leurs bactÃĐries
symbiotiques, qui fixeront l'azote atmosphÃĐrique, ce qui enrichira encore
davantage le sol. Les plantes appelÃĐes à devenir la vÃĐgÃĐtation prÃĐdominante
du lieu s'installent et se rÃĐpandent enfin. Avant que s'ÃĐtablisse une
structure communautaire stationnaire, il faudra peut-Être encore des siÃĻcles,
voire plusieurs millÃĐnaires.

| L'ÃŪle
de Surtsey, au large de la cÃīte mÃĐridionale de l'Islande. - Cette
ÃŪle (prÃĻs d'un kilomÃĻtre carrÃĐ et demi) s'est formÃĐe entre 1963 et
1967, Ã la suite d'une ÃĐruption volcanique. Sa dynamique communautaire
( = ÃĐvolution de la structure et de la composition de la communautÃĐ qui
s'y est constituÃĐe) est ÃĐtudiÃĐe depuis cette ÃĐpoque. Les premiers insectes
volants, apportÃĐs par le vent, y ont ÃĐtÃĐ observÃĐs dÃĻs 1964. Ils ont
fourni une premiÃĻre nourriture à des oiseaux de passage. La suite des
ÃĐvÃĐnements mÃĐtÃĐorologiques a eu pour effet de dÃĐcomposer suffisamment
le substrat pour permettre l'ÃĐtablissement de certaines plantes vasculaires
(les premiÃĻres ont ÃĐtÃĐ observÃĐes dÃĻs 1965) et de lichens nÃĐcessitant
peu de sol. Ces espÃĻces pionniÃĻres (une vingtaine à Surtsey, dont seulement
la moitiÃĐ ont persistÃĐ) ont aidÃĐ Ã ÃĐpaissir une couche de matiÃĻre
organique en dÃĐcomposition, contribuant ainsi à la formation du sol.
Aujourd'hui, environ 70 espÃĻces de plantes poussent sur l'ÃŪle. Des mouettes
y ont ÃĐtabli une colonie sur l'ÃŪle en 1984. Une douzaine d'espÃĻces d'oiseaux
y sont dÃĐsormais rÃĐpertoriÃĐes. Photo : Harvey Barrison,
2016; licence Creative commons. |
Succession
secondaire.
Dans la succession
secondaire, un ÃĐcosystÃĻme est perturbÃĐ gravement, mais des vestiges
de la communautÃĐ prÃĐcÃĐdente subsistent. Une telle succession peut aussi
se produire lorsqu'une communautÃĐ existante a ÃĐtÃĐ supprimÃĐe par une
perturbation qui a cependant laissÃĐ le sol intact. La zone peut retourner
à un ÃĐtat similaire à son ÃĐtat d'origine. Dans le cas d'une zone forestiÃĻre
dÃĐfrichÃĐe pour l'agriculture puis abandonnÃĐe, des plantes herbacÃĐes,
dont les graines ont ÃĐtÃĐ apportÃĐes par le vent ou les animaux, recolonisent
la zone. Ces plantes sont souvent remplacÃĐes par des broussailles
ligneuses, qui elles-mÊmes laisseront la place à des forÊts d'arbres.
Dans le cas, d'un incendie de forÊt, la majeure partie de la vÃĐgÃĐtation
disparaÃŪt et les animaux incapables de fuir sont tuÃĐs. Leurs nutriments,
cependant, sont restituÃĐs au sol sous forme de cendres. De nouveaux individus
pourront ainsi s'installer dans de bonnes conditions. Avant l'incendie,
la vÃĐgÃĐtation ÃĐtait dominÃĐe par de grands arbres ayant accÃĻs à la
principale ressource ÃĐnergÃĐtique vÃĐgÃĐtale : la lumiÃĻre du soleil.
Leur hauteur leur donnait accÃĻs à la lumiÃĻre du soleil tout en ombrageant
le sol et d'autres espÃĻces basses. AprÃĻs l'incendie, cependant, ces arbres
ne sont plus dominants. Ainsi, les premiÃĻres plantes à repousser sont
gÃĐnÃĐralement des plantes annuelles suivies en quelques annÃĐes par des
graminÃĐes
à croissance et propagation rapides et d'autres espÃĻces pionniÃĻres.
Au fil des ans, des espÃĻces, dites espÃĻces intermÃĐdiaires, ÃĐmergeront
: des arbustes, de petits pins, des chÊnes, etc. En un ou deux siÃĻcles,
la forÊt atteindra un ÃĐtat d'ÃĐquilibre stationnaire.
Les populations
Une population correspond
à l'ensemble des individus de la mÊme espÃĻce vivant dans une zone donnÃĐe.
Les membres d'une population dÃĐpendent des mÊmes ressources, sont influencÃĐs
par les mÊmes facteurs environnementaux, et sont susceptibles d'interagir
et de se reproduire entre eux. Les populations peuvent ÃĐvoluer grÃĒce
à l'action de la sÃĐlection naturelle sur les variations hÃĐrÃĐditaires
entre les individus. Les populations fluctuent en fonction de facteurs
biotiques et abiotiques.
CaractÃĐrisation
d'une population.
Effectif
et densitÃĐ d'une population.
Dans un habitat
particulier, une population est caractÃĐrisÃĐe en premier lieu par son
effectif
( = sa taille, le nombre total d'individus qui la composent), et sa densitÃĐ
( = le nombre d'individus dans une zone ou un volume donnÃĐs).
La
variabilitÃĐ gÃĐnÃĐtique à l'intÃĐrieur d'une population et donc le potentiel
d'adaptation de ses membres à l'environnement, explique que les populations
de grand effectif sont souvent plus stables que les populations plus petites.
La
facilitÃĐ d'un individu à trouver un partenaire pour se reproduire peut
dÃĐpendre de la densitÃĐ de la population à laquelle il appartient. A
cause de cela, les populations à faible densitÃĐ peuvent avoir plus de
difficultÃĐs à se maintenir. Par ailleurs, la disponibilitÃĐ des ressources
explique que les petits organismes ont tendance à Être plus densÃĐment
rÃĐpartis que les grands organismes.
SchÃĐmas
de dispersion.
A l'intÃĐrieur d'une
zone gÃĐographique occupÃĐe par une population, sa densitÃĐ moyenne n'est
pas le seul ÃĐlÃĐment à prendre en compte : les densitÃĐs locales
peuvent en effet Être trÃĻs diverses. Les membres de l'espÃĻce peuvent
vivre tous trÃĻs proches les uns des autres ou se trouver ÃĐloignÃĐs, et
lorsqu'ils sont ÃĐloignÃĐs les uns des autres, ils peuvent se distribuer
de façons diffÃĐrentes.
On caractÃĐrise la
relation spatiale qui existe entre les membres d'une population dans un
habitat à un moment donnÃĐ en en dÃĐfinissant le
schÃĐma de dispersion
(ou schÃĐma de distribution). Sa connaissance donne des indications
sur les interactions sociales entre les individus de la population ÃĐtudiÃĐe
et sur la maniÃĻre dont cette population interagit avec son environnement.
Des diffÃĐrences environnementales au niveau local, contribuent aux variations
de la densitÃĐ de population, car certaines zones de l'environnement conviennent
mieux à l'espÃĻce considÃĐrÃĐe.
.
Lorsque les membres
d'une population sont concentrÃĐs en groupes, uniformÃĐment rÃĐpartis
ou dispersÃĐs au hasard, on parle respectivement de de dispersion groupÃĐe,
uniforme et alÃĐatoire.
âĒ
La
dispersion groupÃĐe, qui est le schÃĐma de dispersion le plus souvent
rencontrÃĐ, correspond à la concentrations des membres de la population
dans des zones trÃĻs localisÃĐes. On l'observe chez les plantes qui laissent
tomber leurs graines directement sur le sol ou chez les animaux grÃĐgaires.
L'hÃĐtÃĐrogÃĐnÃĐitÃĐ
de l'habitat peut aussi conduire à une telle dispersion Les plantes
et les champignons se regroupent souvent dans des endroits oÃđ le sol et
d'autres facteurs environnementaux favorisent la germination et la croissance.
Dans les espÃĻces animales, le regroupement peut Être un facilitateur
pour l'accouplement, il peut aussi faciliter l'action de certains prÃĐdateurs
(chasse en meute des loups, par exemple).
âĒLa
dispersion uniforme est le schÃĐma dans lequel les individus d'une
population sont espacÃĐs de maniÃĻre homogÃĻne, gÃĐnÃĐralement avec en
arriÃĻre-plan, une stratÃĐgie d'accÃĻs à des ressources locales limitÃĐes.
On l'observe chez les plantes qui sÃĐcrÃĻtent des substances inhibant la
croissance des individus voisins (allÃĐlopathie nÃĐgative). Des interactions
sociales antagoniques chez certaines espÃĻces animales, comme la dÃĐfense
d'un territoire, peuvent aussi aboutir à une telle dispersion.
âĒ La dispersion
alÃĐatoire, relativement peu commune, correspond à une situation oÃđ
l'espacement des membres d'une population n'est pas prÃĐvisible-:
la position de chaque individu est indÃĐpendante de celle des autres. Ce
schÃĐma de dispersion dÃĐcoule de l'absence de fortes attractions
ou rÃĐpulsions entre les membres de la population ou lorsque les principaux
facteurs physiques ou chimiques pertinents sont relativement homogÃĻnes
dans la zone concernÃĐe. On l'observe, par exemple, avec des
plantes dont les graines, dissÃĐminÃĐes par le vent, germent là oÃđ elles
tombent dans un environnement favorable.
ParamÃĻtres
dÃĐmographiques.
La taille et la
densitÃĐ d'une population dÃĐcrivent une population à un moment donnÃĐ.
Mais lorsqu'on ÃĐtudie une population, il est ÃĐgalement nÃĐcessaire d'en
considÃĐrer l'ÃĐvolution dans le temps. L'effectif et la densitÃĐ d'une
population sont les rÃĐsultantes de divers processus qui lui ajoutent (naissances,
immigration) ou soustraient (dÃĐcÃĻs, ÃĐmigrations) des membres.
Divers paramÃĻtres
permettent de rendre compte de cette ÃĐvolution. On s'en tiendra ici Ã
ne dÃĐfinir que quelques rapports, utiles pour comprendre la dynamique
d'une population.
âĒ
Le
taux de natalitÃĐ est le rapport du nombre de naissances vivantes de
l'annÃĐe à l'effectif total de la population pendant l'annÃĐe. Le rapport
entre les mÃĒles et les femelles ou la structure d'ÃĒge (la proportion
de membres de la population dans des tranches d'ÃĒge spÃĐcifiques), composÃĐe
de nombreux individus en ÃĒge de se reproduire peut augmenter le taux de
natalitÃĐ. Une population nombreuse entraÃŪne un taux de natalitÃĐ plus
ÃĐlevÃĐ parce qu'il y a plus d'individus potentiellement reproducteurs.
âĒ
Le
taux de reproduction est, dans les espÃĻces qui se reproduisent par
voie sexuelle (et donc oÃđ les mÃĒles n'interviennent que pour la distribution
des gÃĻnes), le rapport de leffectif de la progÃĐniture femelle à celui
des mÃĻres.
âĒ Le taux
de fÃĐconditÃĐ. - La fÃĐconditÃĐ est la capacitÃĐ de reproduction d'un
individu au sein d'une population; elle dÃĐcrit le nombre de descendants
qui pourraient Être produits si un individu avait autant de descendants
que possible, en rÃĐpÃĐtant le cycle de reproduction aussitÃīt aprÃĻs la
naissance de la progÃĐniture. En pratique, on dÃĐfinit le taux de fÃĐconditÃĐ,
qui est le rapport du nombre de naissances au cours de la pÃĐriode de rÃĐfÃĐrence
chez les femelles d'un ÃĒge donnÃĐ au moment de la naissance au nombre
de femelles du mÊme ÃĒge pendant cette mÊme pÃĐriode
âĒ Le taux
de mortalitÃĐ est le rapport du nombre de dÃĐcÃĻs de l'annÃĐe à l'effectif
moyen de la population pendant l'annÃĐe. On peut y rattacher le temps qu'un
membre de ce groupe est susceptible de vivre (espÃĐrance de vie). Une grande
taille de population peut entraÃŪner un taux de mortalitÃĐ plus ÃĐlevÃĐ
en raison des maladies, de l'accumulation de dÃĐchets et de la concurrence
dans un milieu aux ressouces limitÃĐes. Dans les populations dÃĐpendantes
de la densitÃĐ, les taux de la mortalitÃĐ augmentent et les taux de natalitÃĐ
diminuent lorsque la densitÃĐ augmente. Dans les populations indÃĐpendantes
de densitÃĐ, ces taux ne changent pas lorsque la densitÃĐ augmente.
Migrations
et mÃĐtapopulations.
Les migrations
peuvent aussi jouer un rÃīle important. Elles peuvent affecter la population
entiÃĻre qui dÃĻs lors change simplement d'habitat (par exemple, la migration
d'un troupeau d'herbivores). La migration peut aussi ne concerner qu'un
nombre dÃĐfini d'individus, ce qui dans ce cas influe sur l'effectif ou
la densitÃĐ d'une population donnÃĐe.
On parle d'ÃĐmigration
lorsque des individus appartenant à une population la quittent pour rejoindre
une zone gÃĐographique diffÃĐrente, et d'immigration lorsque des
individus venus d'une zone diffÃĐrente rejoignent la population considÃĐrÃĐe.
Le solde migratoire (ou accroissement migratoire) est la
diffÃĐrence entre le nombre d'individus entrÃĐes sur un territoire (immigrants)
et le nombre de personnes qui en sont sortis (ÃĐmigrants), au terme
d'un intervalle de temps donnÃĐ. Pour ÃĐvaluer la variation de l'effectif
d'une population (= croissance dÃĐmographique), on l'ajoute à la
diffÃĐrence entre le nombre de naissances (vivantes) dans la population
et le nombre de dÃĐcÃĻs au cours de ce mÊme espace de temps.
Lorsqu'on considÃĻre
ensemble la ou les populations d'origine de migrants et la ou les populations
de destination de ces migrants, on a affaire à une population plus vaste,
dont les composantes sont gÃĐographiquement distinctes, qui prend alors
le nom de mÃĐtapopulation.
StratÃĐgies
d'histoire de vie.
Chaque espÃĻce peut
se caractÃĐriser par l'histoire de vie de ses reprÃĐsentants, autrement
dit, par les caractÃĐristiques propres au mode de reproduction et la survie
des organismes de cette espÃĻce entre leur naissance et leur mort. Les
histoires de vies varient d'une espÃĻce à l'autre parce que chaque espÃĻce,
confrontÃĐe aux limites des ressources (Ã commencer par l'ÃĐnergie) que
peut lui procurer son environnement, a dÃĐveloppÃĐ au cours de l'ÃĐvolution
diverses stratÃĐgies lui permettant d'user des ressources disponibles,
et de les gÃĐrer en les rÃĐpartissant entre les besoins de croissance,
d'entretien et de reproduction, avec la meilleure efficacitÃĐ. Ces stratÃĐgies,
appelÃĐes stratÃĐgie d'histoire de vie, peuvent se rapporter Ã
plusieurs schÃĐmas basÃĐs principalement sur les trois ÃĐlÃĐments suivants
:
âĒ
L'ÃĒge de la premiÃĻre reproduction ou ÃĒge de la maturitÃĐ. C'est le moment
oÃđ la reproduction commence. Il existe une relation entre cet ÃĒge et
l'engagement dans les soins parentaux. Les individus d'une espÃĻce peuvent
se reproduire tÃīt dans leur vie pour s'assurer de survivre jusqu'Ã l'ÃĒge
de la reproduction ou se reproduire plus tard dans leur vie pour devenir
plus grands et en bonne santÃĐ et Être mieux à mÊme de prodiguer des
soins parentaux.
âĒ La frÃĐquence
de reproduction des organismes. Les individus d'une espÃĻce peuvent se
reproduire une fois (semelparitÃĐ) ou plusieurs fois (itÃĐroparitÃĐ) au
cours de leur vie. Le facteur dÃĐterminant est ici le taux de survie de
la progÃĐniture.
+ La sÃĐmelparitÃĐ
se produit lorsque les individus d'une espÃĻce ne se reproduisent qu'une
seule fois au cours de sa vie, puis meurent. Les espÃĻces sÃĐmelpares utilisent
la majeure partie de leur budget de ressources au cours d'un seul ÃĐvÃĐnement
reproductif, sacrifiant leur santÃĐ au point qu'elles ne survivent pas.
La sÃĐmelparitÃĐ s'observe souvent chez des espÃĻces vivant dans des environnements
trÃĻs variables ou imprÃĐvisibles.
+L'itÃĐroparitÃĐ
dÃĐcrit les espÃĻces dont les reprÃĐsentants se reproduisent à plusieurs
reprises au cours de leur vie. Certains animaux ne peuvent s'accoupler
qu'une fois par an, mais survivent à de multiples saisons d'accouplement.
L'itÃĐroparitÃĐ caractÃĐrise surtout des espÃĻces vivant dans un environnement
stable
âĒ Le nombre
de descendants produits dans chaque ÃĐpisode de reproduction (nombre de
graines produites par la plante, taille de la portÃĐe ou de la couvÃĐe
chez les animaux). Le nombre de descendants a souvent un lien avec la frÃĐquence
de reproduction : la descendance est nombreuse dans les environnements
incertains, ce qui augmente la probabilitÃĐ qu'au moins certains individus
survivront. Dans des environnements plus sÃŧrs, quelques petits, disposant
de provisions suffisantes, auront plus de chances de survivre jusqu'Ã
l'ÃĒge de la reproduction. Il existe aussi une relation inverse entre
la fÃĐconditÃĐ et les soins parentaux, qui sont plus importants quand la
progÃĐniture est peu nombreuse.
Les modÃĻles de la
croissance dÃĐmographique.
Des modÃĻles mathÃĐmatiques
peuvent Être utilisÃĐs pour dÃĐcrire les changements survenant dans une
population ou pour mieux les prÃĐdire. Deux modÃĻles simples et idÃĐalisÃĐs
sont prÃĐsentÃĐs ici. Le premier montre que les populations disposant de
ressources illimitÃĐes connaissent une croissance exponentielle, avec un
taux de croissance accÃĐlÃĐrÃĐ. Dans le second modÃĻle, on pose l'existence
d'une limite au ressources disponibles et dans ce cas, la croissance de
la population, d'abord rapide, se ralentit puis se stabilise à un niveau
maximal. Les situations rÃĐelles appellent cependant des approches plus
complexes.
Croissance
exponentielle.
Si l'on ne prend
pas en compte les effets des migrations, Ã chaque gÃĐnÃĐration l'effectif
d'une population varie en fonction de son taux de natalitÃĐ
n et
de sonb taux mortalitÃĐ m. Le taux
r de croissance intrinsÃĻque
de cette population s'ÃĐcrit alors : r = n - m. La valeur de r
peut Être positive (augmentation de l'effectif de la population); ou nÃĐgative
(diminution de l'effectif de la population; croissance dÃĐmographique nÃĐgative),
ou nulle (pas de changement dans la taille de la population; croissance
nulle). Dans le cas d'une croissance nÃĐgative qui se maintient, la population
est appelÃĐe à s'ÃĐteindre.
Le taux de croissance
d'une population, c'est-Ã -dire la variation de son effectif N en
un temps t donnÃĐ est ÃĐgal à son taux croissance intrinsÃĻque
r
multipliÃĐ par cet effectif. En temps continu, on peut exprimer cela sous
la forme d'une ÃĐquation diffÃĐrentielle :
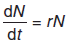
Cette ÃĐquation signifie
que le taux de variation de la population N Ã un instant t
(dÃĐrivÃĐe de N par rapport au temps) est ÃĐgal au produit du taux
de variation intrinsÃĻque r par l'effectif de la population. Lorsqu'on
l'intÃĻgre, on obtient un effectif de la population ÃĐvoluant en fonction
du temps de façon exponentielle.
Croissance
logistique.
Dans le monde rÃĐel,
les ressources ne sont pas illimitÃĐes et la croissance exponentielle ne
peut exister (et encore de maniÃĻre approximative) que tant que les besoins
de la population restent nÃĐgligeables par rapport aux ressources
disponibles. A partir d'un certain moment, la croissance de la population
est visiblement freinÃĐe par l'existence de cette limite : chaque individu
a accÃĻs à une quantitÃĐ dÃĐcroissante de ressources et il existe une
limite à la quantitÃĐ d'individus qui peuvent vivrent dans un habitat
donnÃĐ. Un modÃĻle plus rÃĐaliste doit incorporer la notion de capacitÃĐ
de charge (K), qui est la valeur maximale que peut atteindre une
population dans un environnement donnÃĐ. Le nombre d'individus pouvant
s'ajouter à une population à un moment donnÃĐ est K-N. Le rapport
(K-N)/K
correspond alors à la fraction de la capacitÃĐ de charge disponible pour
une croissance supplÃĐmentaire. L'ÃĐquation prÃĐcÃĐdente doit Être modifiÃĐe
pour rendre compte de cette contrainte. Elle devient :
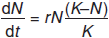
L'intÃĐgration de l'ÃĐquation permet de tracer
une courbe reprÃĐsentative de N en fonction du temps qui prend la forme
d'un sigmoÃŊde, c'est-Ã -dire d'une courbe en forme de S. La droite d'ÃĐquation
y = K est son asymptote quand t tend vers l'infini
(Les fonctions).
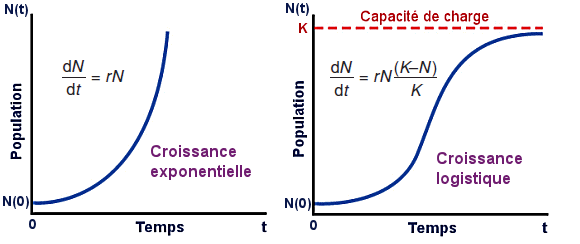
| Croissance
exponentielle et croissance logistique. - L'ÃĐquation de la croissance
exponentielle reprÃĐsente la croissance potentielle d'une population dans
un environnement aux ressources illimitÃĐes. Ce modÃĻle prÃĐdit que plus
une population est grande, plus elle grandit rapidement (rÃĐtroaction positive).
L'ÃĐquation de la croissance logistique prend en compte l'existence d'une
limite des ressources disponibles. Les premiers temps de la croissance
d'une population soumise à telle limitation ressemblent à ceux de la
croissance sans limitation de ressources, mais l'existence d'une capacitÃĐ
de charge se fait vite sentir (rÃĐtroaction nÃĐgative); le taux de croissance
diminue à mesure que la population approche de sa capacitÃĐ de charge.
La population se stabilise prÃĻs de cette valeur limite. |
Croissance
logistique et population rÃĐelle.
Plus rÃĐaliste que
le modÃĻle exponentiel, le modÃĻle logistique de la croissance d'une population
n'en est pas moins une approximation des situations rÃĐelles, et il peut
exister bien d'autres maniÃĻres pour l'effectif d'une population d'ÃĐvoluer
(cycliques comme dans l'exemple LiÃĻvres-Lynx, ci-dessous; chaotiques;
etc.). Plusieurs facteurs jouent dans l'ÃĐvolution de l'effectif d'une
population rÃĐelle. On a dÃĐjà ÃĐvoquÃĐ le rÃīle des migrations (une ÃĐmigration
peut intervenir quand les ressources sont plus difficiles d'accÃĻs). On
doit aussi tenir compte de la compÃĐtition intraspÃĐcifique : ÂŦ tous les
animaux sont ÃĐgaux, mais certains sont plus ÃĐgaux que d'autres Âŧ (G.
Orwell, 1945) pour accÃĐder aux ressources (cela vaut aussi pour les populations
vÃĐgÃĐtales, bactÃĐriennes, etc.). La densification de la population, qui
peut attirer davantage de prÃĐdateurs, et l'accumulation de dÃĐchets
peuvent aussi Être des freins à l'accroissement dÃĐmographique.
Par ailleurs, le
modÃĻle logistique suppose que les espÃĻces s'adaptent instantanÃĐment
à la croissance et à l'ÃĐvolution de la population et se rapprochent
en douceur de la capacitÃĐ de charge. Toutefois, dans la plupart
des populations rÃĐelles, il s'ÃĐcoule un certain temps avant que les facteurs
limitatifs nÃĐgatifs de la croissance dÃĐmographique ne deviennent apparents.
Si, par exemple, la disponibilitÃĐ de la nourriture commence à diminuer
dans une population, la reproduction diminue ÃĐgalement. Cependant, le
taux de natalitÃĐ peut ne pas Être rÃĐduit immÃĐdiatement car les femelles
peuvent utiliser leurs rÃĐserves d'ÃĐnergie pour continuer à se reproduire
pendant une courte pÃĐriode. Cela signifie que dans certains cas l'espÃĻce
est en mesure de dÃĐpasser sa capacitÃĐ de charge avant d'atteindre une
densitÃĐ relativement stable. La capacitÃĐ de charge peut aussi se modifier
pour des raisons extÃĐrieures à la population, ou simplement parce que
les ressources dans lesquelles puise la population ne se renouvellent plus.
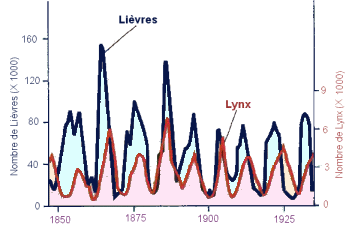
| Cycles
de populations engendrÃĐs par une interaction proie-prÃĐdateur. - Cet
exemple est tirÃĐ de l'ÃĐtude de l'ÃĐvolution des populations de LiÃĻvres
amÃĐricains et de Lynx dans le Nord de l'Ontario pendant un siÃĻcle, Ã
partir du nombre de peaux vendues par les chasseur de la Compagnie
de la baie d'Hudson. Lorsque le nombre de liÃĻvres augmente, les lynx
disposent de plus de nourriture et leur nombre augmente. Il s'ensuit que
le nombre de liÃĻvres diminue de nouveau. La nourriture venant à manquer
aux lynx, leur population diminue à son tour. Moins chassÃĐs, les liÃĻvres
peuvent de nouveau se multiplier : le cycle recommence. |
 |
En
librairie. - Collectif, Dictionnaire
de l'ÃĐcologie,
Encyclopaedia universalis, 2001; Dieter Heinrich et Manfred Hergt, Atlas
de l'ÃĐcologie, Livre de Poche, 1993;
Jean-Paul DelÃĐage, La BiosphÃĻre :
Notre terre vivante, Gallimard, 2001;
Robert Barbault, Ãcologie gÃĐnÃĐrale,
Structure et fonctionnement de la biosphÃĻre: Structure et fonctionnement
de la biosphÃĻre, Dunod, 2021; Jacques
Grinevald, La BiosphÃĻre de l'AnthropocÃĻne
: Climat et pÃĐtrole, la double menace,
Georg Editeur, 2008.
Alain PavÃĐ, Comprendre la biodiversitÃĐ.
Vrais problÃĻmes et idÃĐes fausses: Vrais problÃĻmes et idÃĐes fausses,
Seuil, 2019; Christian LevÊque, La
biodiversitÃĐ : avec ou sans l'homme? RÃĐflexions d'un ÃĐcologue sur la
protection de la nature en France,
QUAE GIE, 2017; Pierre-Henri Gouyon, CÃĐdric Gaucherel, Jean-Louis
Dessalles, Le Fil de la Vie. La face
immatÃĐrielle du vivant, Odile Jacob,
2016.
Serge Frontier et Denise Pichod-Viale et al, ÃcosystÃĻmes,
Structure, Fonctionnement, Ãvolution,
Dunod, 2008; François BrÃĐchignac et Lisa Cauvin, L'ÃĐcosystÃĻme:
La dimension nÃĐgligÃĐe du vivant,
L'Harmattan, 2022.
Neil Campbell, Jane Reece et al., Biology,
2005; Eugene Odum, Ecology,
1963-1975; P. H. Collin, Dictionary
of environment and ecology, 1992-2004. |
|
|
